


 回忆像条狗6666
回忆像条狗6666
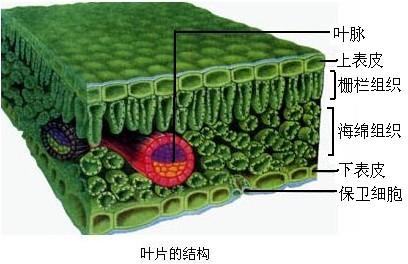
范文一:下图1表示绿色植物叶肉细胞部分结构中的某些生命活动过程
下图1表示绿色植物叶肉细胞部分结构中的某些生命活动过程,①-⑦代表各种物质,甲、乙代表两种细胞器。图2表示在温度、水分都适宜时,该植物叶片CO2释放量随光照强度变化的曲线。请据图回答下列问题:
(1)图1中,甲、乙两种结构的名称依次是;甲、乙两种细胞器的膜面积都很大,其中乙增加膜面积主要是通过 。
(2)图1中①的名称是 ,⑦的名称是 。
(3)若该绿色植物长时间处于黑暗状态中,则图1中“⑥→⑤→⑥”的循环能否进行?。
(4)与图2 中的A点相对应时,图1中能产生ATP的细胞器有 ;当光照强度达到图2中的 D点以后,限制光合作用强度继续上升的主要环境因素
是 。
共9分,每空1分)
(1
)叶绿体 线粒体 内膜向内腔折叠形成嵴
(2)水 O2
(3)不能 缺少光反应提供的[H]和ATP
(4)乙(线粒体) CO2浓度
范文二:喷施草甘膦后叶肉细胞的扫描电镜观察
喷施草甘膦后叶肉细胞的扫描电镜观察
第29卷第2期
2011年6月
湖北民族学院(自然科学版)
JournalofHubeiUniversityforNationalities(NaturalScienceEdition)
Vo1.29No.2
Jun.2011
喷施草甘膦后叶肉细胞的扫描电镜观察
卜贵军,郑小江,崔琳,王学东
(1.生物资源保护与利用湖北省重点实验室(湖北民族学院),湖北恩施445000;
2.湖北民族学院生物科学与技术学院,湖北恩施445000;
3.东北农业大学生命中心,黑龙江哈尔滨150030)
摘要:以东农42为材料,对处于营养生长期的东农42喷施草甘膦,从叶肉细胞形态
结构方面来研究东农42的
作用.利用扫描电镜,研究喷施草甘膦后大豆东农42叶肉细胞的栅栏组织和海绵
组织的变化,来从微观上揭示草
甘膦对大豆的伤害.研究结果表明东农42的叶肉细胞在喷施草甘膦后栅栏组织和
海绵组织的细胞皱缩,变形.我
们尽可能对其原因进行探究,为草甘膦作用机理研究起到一定的作用.
关键词:草甘膦;大豆;栅栏组织;海绵组织
中图分类号:Q944.56文献标识码:A文章编号:1008—8423【2011)O2—0220一o3
GlyphosateEffectsonUltrastructureChangeofSoybean (GlycinemaxL.)MesophyllCells
BUGui—jun,ZHENGXiao—jiang,CUILin,WANGXue—dong3
(1.KeyLaboratoryofBiologicResourcesProtectionandUtilizationofHubeiPronvince,
HubeiUniversityforNationalities,Enshi445000,China; 2.SchoolofBiologicalScienceandTechnology,HubeiUniversityforNationalities,EnsH44
5000,China;
3.LifeCenterofNoahe~tAgricultureUniversity,Harbin150030,China) Abstract:Inthisstudy,wechooseDongnong42forthemateria1.Byusingscanningelectronm
icroscope
(SEM),westudypalisadetissueandspongytissuemicrostructuralchangesinphotosynthesis
organsoy—
beanDongnong42,inresponsetoglyphosate.TheresultsshowedthatsoybeanDongnong42
Mesophyll
Cellswerethomughlydestroyedbyglyphosate.Inaddi6on,wetryourbesttoprobeitsreasons.
Itmaybe
usefulforthestudyofglyphosatemechanism.
Keywords:glyphosate;soybean;palisadetissue;spongytissue 草甘膦又叫农达,见绿杀,镇草宁,是一种广谱性的,内吸传导型除草剂】J.草甘膦经
叶片表面的角质
膜和气孔吸收后,在植物细胞中,被转化为水溶性的盐,它的运输的主要方式是,从
韧皮部随着光合作用的产
物,输送到生长旺盛的部位,属于由"源"向"库"的运输.由于是传导性除草剂,它可
以杀死整个植株,其中也
包括根,从而可以预防由根引起的再生,达到斩草除根的功效.
草甘膦的化学结构与磷酸烯醇式丙酮酸(PEP)的类似,这就使得草甘膦能与PEP
竞争性抑制合酶的活
性,形成EPSPS合酶一3一磷酸莽草酸一草甘膦的复合物,从而抑制了合成EPSP
的EPSP合成酶J,阻断了莽
草酸途径和芳香族氨基酸的合成.草甘膦阻断芳香族氨基酸和一些芳香化合物的
生物合成,就会扰乱生物体
正常的氮代谢而使植物体死亡.SuttonRF等的研究表明低浓度的草甘膦可能比高
浓度的更有效J.这是由
收稿日期:2011—03一O1.
基金项目:黑龙江省自然科学基金项目(C201023).
作者简介:贵军(1981一),男,硕士,主要从事植物结构生物学的研究;通讯作者:王学东(1958一),男,研究员,主要从事植物结构生物学的
研究.
第2期卜贵军等:喷施草甘膦后叶肉细胞的扫描电镜观察22l 于高浓度的草甘膦一接触植物就可以杀死组织细胞,而不能使草甘膦得到传递.而低浓度的草甘膦却可以在
体内转运,直到达到根部和其他部位,杀死整个植株.
正是由于草甘膦的诸多优点,使得草甘膦的应用极为广泛.在施用草甘膦时杀草时,就可能导致草甘膦
飘逸至农作物上,因此,有必要研究草甘膦对农作物的伤害.此前的研究一般都利用透射电镜来研究草甘膦
对叶肉细胞的破坏】,用扫描电镜来研究草甘膦对表皮细胞的破坏?引.本研究利用扫描电镜,观察喷施草甘
膦后大豆叶肉细胞的变化,比利用透射电镜更具立体感.
1材料与方法
1.1材料
大豆东农42(Glycinm眦L.Dongnong42).
1.2大豆叶片超微结构的观察
本试验年在东北农业大学农学试验站进行,行距65cm,行长35m,株距10cm. 在幼荚期,天气晴好时,对实验区部分东农42的叶片施用草甘膦.草甘膦浓度按说明书比例配制{3:
400配药,由鹤壁市农林制药有限公司生产的草甘膦异丙胺盐(Glyphosate),剂型:水剂含量:41%}.分别在
第1次喷药前(0d)和喷药后的第1—7d的上午9:00,11:00,取第一叶脉与第二叶脉之间的部分,用双面刀
片,切成宽度约为2mm,长约5mm的长条用于制备扫描电镜样.切好后将两部分用牙签轻轻拨入盛有用磷
酸缓冲液配制的pH为6.8的戊二醛固定液的小瓶中,然后置于冰盒中. 扫描电子显微镜样品制备:将取回的样品经过固定后,在经过冲洗,脱水,置换,.用ES-2030(HITACHI)
型冷冻干燥仪对样品进行干燥,粘样,镀膜:用E—1010(HITACHI)型离子溅射镀膜仪在样品表面镀上一层
15nm后厚的金属膜(金或铂膜).将处理好的样品放入样品盒式中待检.最后在日立公司的S一3400扫描电
镜下观察,并拍照.
2实验结果
2.1大豆叶片栅栏组织细胞的扫描电镜观察
东农42的在没喷施草甘膦前,栅栏组织细胞,呈柱状,排列紧密,细胞非常规则(图版l一1);喷药后第1
天和第2天,栅栏组织细胞未见明显变化(图版I一2,3);喷药后第3天,栅栏组织细胞,出现变形,细胞变得
不规则,有皱缩(图版I一4);喷药后4d,栅栏组织细胞变形程度加大,细胞体积急剧减小,皱缩程度加剧(图
版I一5);喷药后第5天以后,栅栏组织细胞变形非常严重,基本被瓦解(图版I一6,7,8).
2.2大豆叶片海绵组织细胞的扫描电镜观察
东农42的在没喷施草甘膦前,海绵组织细胞,非常规则(图版I一1);喷药后第1天没有明显变化(图版
I一2);喷药后第2天,海绵组织细胞有一定皱缩,变形(图版I一3);喷药后第3天,海绵组织细胞,变形明
显,皱缩加剧,细胞变得不规则(图版I一4);喷药后4天,海绵组织细胞变形程度加大,细胞体积急剧减小,
皱缩加剧(图版I一5);喷药后第5天以后,海绵组织细胞变形非常严重,基本被瓦解(图版I一6,7,8).
3结果分析与讨论
本试验利用扫描电镜,来研究草甘膦对东农42的叶片的栅栏组织和海绵组织的影响,结果发现,在按照
说明书的浓度喷施草甘膦的情况下,栅栏组织和海绵组织在一定时问内对于草甘膦的胁迫都会有一定的伤
害,而且随着天数的增加,伤害程度加大.研究发现海绵组织出现变形的时间比栅栏组织的稍早,这可能是由
于草甘膦通过气孔进人叶片的速度要大于通过角质层进入的速度.栅栏组织在接近叶片的上表皮处,海绵组
织在接近叶片的下表皮处,而上表皮的气孔数要少于下表皮的气孑L数,因此从下表皮,也就是靠海绵组织一
侧进入的草甘膦要比栅栏组织一侧要多,因此伤害也就越快.
草甘膦进人大豆东农42叶片后,通过细胞壁,细胞膜进入叶肉细胞,在叶肉细胞的叶绿体中,能够竞争
抑制EPSPS的作用,从而使得东农42的芳香族氨基酸合成受阻,影响到含有芳香族氨基酸的蛋白质合成,
以及芳香族氨基酸及其衍生物的代谢,由于草甘膦作用位置在叶绿体中,所以会导致细胞内部叶绿体变形,
另外,由于草甘膦作为一种外界物理作用,可能会由于渗透压等的原因引起液泡等结构变化,从而在微观结
222湖北民族学院(自然科学版)第29卷
图1喷施草甘膦后东农42叶片的栅栏组织和海绵组织的扫描电镜观察 图版1—1未喷施草甘膦的东农42的叶片的栅栏组织和海绵组织细胞, 图版1—2,3,4,5,6,7,8为喷施草甘膦1,7d东农42的叶片的栅栏组织和海绵组织的变化.
Fig.1Dongnong42leavesPahsadetissueandspongytissuecellschangeafterspraying
glyphosateunderscanningelectronmicroscopy
Fig.1-1Dongnong42leavesPalisadetissueandspongytissuecellswithnospraying,
Fig.1-2.3,4,5,6,7,8Dongnong42leavesPalisadetissueandspongytissuecellschangeat1—7daysafterspraying.
构上表现出来.这与本人对于草甘膦对大豆叶片的透射电镜观察结果是一致的J. 总之,草甘膦对叶肉细胞的所产生的伤害,可能是一系列综合因素的结果,一开始是由渗透压,以及对芳
香族氨基酸合成抑制所造成的,以后是由于芳香族氨基酸及其类似物缺乏,影响蛋
白质合成及代谢,使细胞
内代谢崩溃所致.
参考文献:
[1]MalikJ,BarryG,KishoreG.Minireview:Theherbicideglyphosate[J].Biofacrors,1989,2:17—25.
[2]BradshawLD,PadgetteSR,KimballSL,eta1.Perspectivesonglyphosateresistance[j].WeedTechnol,1997,11(1):189—198.
[3]KennedyER,JordanPA.Glyphosateand2,4一
D:theimpactoftwoherbicidesonmoosebrowseinforestplantations[J].Alces,1985,1:1—
11.
[4]SammonsRD,GruysKJ,AndersonKS,eta1.Reevaluatingglyphosate8satransitionstateinhibitorofEPSPsynthase:identificationofanEPSP
[5]
6]
7]
8]
9]
l0
synthaseEPSPglyphosateternarycomplex[J].Biochem,1995,34(19):6433—6440.
MedowellLM,KlugCA.BeusenDD.Ligandoftheternarycomplexof5一
enolpyrevylshikimate-3一phosphatesynthasefromrotational-echo
double—resonanceNMRfJ].Biochem,1996,35(17):5395—5403.
SarohaM.K.SridharP,MalikVS.Glyphosate-tolerantcrops:Genesandenzymes[J].JPlantBiochemBiotechnol,1998,7:65-772.
TanS,EvansR,SinghB.Herbicidalinhibitorsofaminoacidbiosynthesisandherbicide-tolerantcrops[J].AminoAcids,2006,30:195-204.
SuttonRF.Glyphosateherbicide:anassessmentofforestrypotential[J].ForChronicle,1978,54(1):24—28.
贵军,郑小江,王学东,等.草甘膦对大豆叶片超微结构及生化指标的影响[J].中国油
料作物,2010,32(2):285—289.
]贵军.喷施草甘膦对大豆表皮细胞超微结构影响的研究[J].湖北民族学院:自然科
学版,2011,29(1):15—18.
范文三:解决光下叶肉细胞气体转移问题的关键
解决光下叶肉细胞气体转移问题的关键
不同光照条件下气体在叶绿体和线粒体之间以及叶肉细胞和空
气之间的交换,一直是困扰老师和学生的大问题,解决这一问题的
关键就是自由扩散。
1.自由扩散
人教版教材是这样给出的自由扩散:物质通过简单的扩散作用进
出细胞叫作自由扩散。像co2和o2这样的物质通过细胞膜进出细
胞就是自由扩散,细胞膜、线粒体膜、叶绿体膜都是生物膜,co2
和o2跨膜进出线粒体、叶绿体也是自由扩散。自由扩散的重要特
点就是顺浓度梯度扩散,物质运输的速度主要取决于浓度差。
co2和o2在空气、叶绿体和线粒体中的浓度是怎样的呢?叶绿体
在光下能不断进行光合作用消耗co2生成o2,可认为co2浓度最低,
o2浓度最高;线粒体不断进行有氧呼吸消耗o2生成co2,可认为
o2浓度最低,co2浓度最高;空气中co2和o2浓度介于叶绿体中
的浓度和线粒体中的浓度之间。即光下三者之间o2浓度的关系:
叶绿体中o2浓度>空气中o2浓度>线粒体中o2浓度;光下三者
之间co2浓度的关系:线粒体中co2浓度>空气中co2浓度>叶绿
体中co2浓度。叶绿体和线粒体之间的o2浓度差最大;线粒体和
叶绿体之间的co2浓度差最大。
2.气体转移
在光下,自然环境中,气体的转移情况如图所示。
3.分析不同光照条件下的气体转移
根据如图所示的不同光照条件下光合作用强度曲线(用co2的吸
收和释放来表示),克服气体进出平衡的那一部分,我们可以这么
理解:
当光照强度为零时,也就是a点对应的位置。只有呼吸作用而没
有光合作用,线粒体沿d途径从空气中获取o2,产生的co2沿c途
径排入空气。
当光照强度处于ob段,即光合作用强度小于呼吸作用的强度时。
对于线粒体来说,o2可来源于叶绿体和空气,考虑到叶绿体和线粒
体之间o2的浓度差最大,可以说o2快速来源于叶绿体,不足的由
空气中的o2补充,即o2主要沿b、d途径进入线粒体;同理分析线
粒体产生的co2主要沿 a、c途径扩散到叶绿体和空气。对于叶绿
体来说,叶绿体可快速从线粒体处获取co2,由于光合作用强度很
弱,所需的co2完全可由线粒体提供,即叶绿体所需的co2主要沿
a途径进入;同理分析叶绿体产生的少量o2主要沿b途径扩散。
当光照强度为b,即光合作用强度和呼吸作用强度相等时。对于线
粒体来说,考虑到叶绿体和线粒体之间o2的浓度差最大,可以说
o2快速来源于叶绿体,且叶绿体产生的o2恰好满足线粒体的需求,
即线粒体所需的o2主要沿b途径进入;同理分析线粒体产生的co2
可沿a途径散失。对于叶绿体来说也是如此分析,叶绿体所需的co2
主要沿a途径进入, 产生的o2主要沿b途径散失。
当光照强度大于b,光合作用强度大于呼吸作用强度时。对于线粒
体来说,所需的o2可快速来源叶绿体,由于呼吸作用相对较弱,
需要的o2较少,就是说线粒体需要的o2沿b途径进入就能满足, 产
生的co2的主要沿a途径散失;对于叶绿体来说,所需的co2可快
速来源于线粒体,不能满足的部分由空气中的co2补充,即co2主
要沿a、e途径进入叶绿体,叶绿体产生的o2主要沿b、f途径散
失。
同样的分析方法,请看下面的例题,注意叶片周围的空气是经过
naoh溶液的。
4.例题
用大豆新鲜绿叶进行如图所示的实验,置于充足的光照下,分析d
处的气体成分,最可能的是( )
a.氧气的量多于b处 b.氧气的量与b处相等
c.没有co2和o2 d.co2的量多于b处
分析:经过naoh溶液后,b处没有co2,浓度为零;叶绿体不断
消耗co2认为其浓度极低接近为零;只有线粒体进行呼吸产生co2,
认为其浓度极高。现在co2的浓度差情况是这样的:线粒体处的co2
浓度与b处”空气“ 的co2浓度差和线粒体处的co2浓度与叶绿
体处的co2浓度差相等。根据影响自由扩散的因素可知,线粒体产
生的co2扩散进入叶绿体和扩散进入空气的几率是一样的,进入空
气的co2随气流就到了d处,结果就是d处有co2而b处无co2,
由于叶肉细胞释放出co2,也可分析出d处o2浓度少于b处。
参考答案:d
立足基础,从自由扩散的基本概念与实质入手是解决光下气体转
移问题的关键,也是学生学好理科知识的关键。
范文四:盐害对淹水胁迫下水稻叶肉细胞的影响
盐害对淹水胁迫下水稻叶肉细胞的影响
纪群 戴盛桃 祝天慧 司晶 张磊
摘要: 本实验通过有高浓度的离子培养液做培养基,观察盐害对水稻淹水胁迫的影响。我们主要观察细胞整体形态和叶绿体,线粒体的形态,测定 MDA (丙二醛)、过氧化氢酶和超氧歧化酶活性来反映整个细胞的活性。实验发现,经过盐害的水稻与对照组相比,无论
从细胞形态还是生理指标看,抗淹能力都下降明显。 关键词:盐害 淹水胁迫 形态 生理
受淹的危害主要是造成受淹植物组织出现缺氧。很多研究者通过对根细胞的研究,对在淹水所致的完全缺氧条件下植物细胞出现死亡的原因 , 提出许多不同的解释。越来越多的研究者认为 , 是无氧发酵所产生的能量不能充分满足植物细胞的需要 , 引起植物细胞内各种细胞器结构的破坏 , 最终导致植物细胞死亡。要提高植物的抗淹能力,关键是提高其抗缺氧的能力 4 。前的这方面的研究主要停留在对受淹植物某些生理指标的检测,而对其细胞结构,细胞器的变化缺乏深入研究,关于盐害对受淹植物的影响更是缺乏研究 2 。植物在受淹的过程中,叶肉细胞的很多细胞结构都回发生明显的变化。土壤淹水或过湿是一种十分常见的自然环境。根据联合国粮农组织和国际土壤协会绘制的世界地图估算 , 全球约有 12% 的土地处于常年过湿状态。淹水胁迫常常影响植物生长发育 , 过多的水分阻碍了植物与大气环境间的气体交换 , 造成植物受淹组织缺氧。由于高等植物均为需氧生物 , 必须从环境中获取氧气 , 以维持其正常的生理代谢及生长发育。因此 , 淹水胁迫往往会导致作物严重减产。在长期的进化过程中 , 一些植物如水稻、油菜等已经演化出了一套适应机制和
策略。而另外一些重要农作物如烟草、小麦、玉米等对淹水非常敏感 , 缺乏有效的耐淹防御机制。因此 , 阐明淹水对植物生长发育造成危害的根本原因以及植物相应的耐淹机制 , 对于发展农业生产具有十分重要的现实意义。
1. 材料和方法
1.1 实验材料:“粤香粘”水稻,由广东农业科学研究所提供。
1.2 实验方法
水稻种子于 2004 年 11 月 24 日开始播种于放有棉花的培养皿中,添加以下培养液。 表一 各实验组所用的培养液
正常组 盐害组 1 盐害组 2 盐害组 3 离子浓度 Hoagland 溶液Hoagland 溶液 Hoagland 溶液 Hoagland 溶液
的 1/10
从 11 月 24 日到 12 月 7 日盐害组一直浇 Hoagland 溶液,从 12.7 , 12.14 盐害组也浇 1/10 浓度的 Hoagland 溶液。对照组一直浇 1/10 浓度的 Hoagland 溶液。 至 12.14 开始进行水淹,将水稻全部浸没,连续 3 天于上午 8 : 30 取叶片进行实验。 用机械法分离叶肉细胞,然后分别进行液泡、线粒体染色观察。具体方法见《细胞生物学实验教程》(王金发、何炎明) P44 。
叶肉细胞生理指标的测定:用紫光吸收法测定淹水 1 、 2 、 3 天后各组叶肉细胞中丙二醛( MDA )、过氧化氢酶( POD )、超氧物歧化酶( SOD )活性。
2 实验结果
2.1 经过 20 天的培养,干旱组做三组。培养的水稻幼苗如下:
图 1 对照组(左)与两周盐害组(右)形态整体
经过 13 天盐害培养的水稻幼苗与对照组相比,盐害组叶片有些泛黄。长势也没有对照组好。
2.2 叶肉细胞形态观察
从以下各组的对照图片可以看出:
淹水 3 天后,叶肉细胞整体看来逐渐模糊,,细胞边界变得不明显。 淹水 2 天后 , 液泡膜发生明显内陷。淹水 3 ,后 , 液泡膜内陷加剧 , 呈极度松弛状态 ; 叶绿体被膜局部向外突出一个由单层膜包裹的泡状结构。淹水 2 天叶绿体被膜破坏加剧 , 成为一松弛的单膜结构 , 同时 , 基质类囊体出现空泡化。淹水 3 天后 , 叶绿体的破坏进一步加剧 : 被膜完全消失 , 基质类囊体开始解体。
A B
C D
F E
F G
( 100 × 4 )
H G
H I
图 2 淹水前后细胞形态结构的变化,其中 A 、 B 为对照组淹水前后细胞形态变化, C-G 为盐害组淹水前后细胞形态变化。 H 、 G 为淹水后液泡染色观察; H 、 I 为淹水后线粒体染色观察。
2.3 淹水对水稻叶肉细胞生理指标的影响
2.3.1 . 丙二醛( MDA )含量变化
图 3 所示,在淹水的过程中丙二醛的含量急剧上升。在未淹之前,盐害组叶肉细胞中 MDA 的含量略高于对照组,这可能是由于盐害造成的干旱,使得盐害组细胞中 MDA 的含量升高。在淹水的过程中,三个盐害组 MDA 的含量始终高出对照组,其 MDA 含量的变化率稍微高于对照组,由于丙二醛是植物缺氧呼吸产生的有害物质,其积累对植物细胞有毒害作用。因此单从 MDA 的积累变化来看,盐害会降低水稻的抗淹能力。
表 2 水淹对水稻叶肉细胞中 MDA 含量的影响
淹水时间( D ) 对照组 盐害组 1 盐害组 2 盐害组 3 平均值 0 0.21364 0.24233 0.2953 0.25266 0.39099
1 0.34244 0.4417 0.34854 0.4848 0.49183
2 0.48166 0.62232 0.57262 0.62275 0.4417
3 0.6082 0.7621 0.89364 0.69607 0.69654
A
B
图 4 淹水过程中叶肉细胞中 MD 含量的变化 (A 曲线图, B 方柱图 ) A 图曲线 1 为对照组, 5 为盐害组的平均值变化。 B 图蓝色为对照组,褐色为盐害组平均值。
2.POD 酶活性测定
图 4 所示,淹 1 天后, POD 活性急剧下降。其中对照组下降最明显。未淹水前,对照组 POD 酶活性显著高于三个盐害组。淹水 2 、 3 后各组 POD 酶活性有所上升,稍低于淹水前酶的活性。这可能是因为水稻比较耐淹,连续淹水 3 天对于其细胞内酶活性影响不大。没有像预期那样下降。从两组的对照来看,盐害组细胞内 POD 酶活性在水淹前后都显著低于对照组。
表 3 淹水过程中叶肉细胞中过氧化氢酶( POD )活性的变化
淹水时间
( D ) 对照组 盐害组 1 盐害组 2 盐害组 3 平均值
0 155.78 51.93 16.17 31.53 33.21
1 31.06 13.49 4.94 16.14 11.52
24.77 20.57 25.65 23.67 2 133.57
3 178.15 40.67 26.7 46.14 37.84
A
B
图 5 :淹水过程中叶肉细胞中过氧化氢酶( POD )活性的变化 (A 曲线图, B 方柱图 ) A 图曲线 1 为对照组, 5 为盐害组的平均值变化。 B 图蓝色为对照组,褐色为盐害组平均值。
3.SOD 酶活性测定
超氧物歧化酶,是植物体内清除自由基的一种重要的酶。图 5 显示对照组的的趋势是先下降然后上升。这是由于淹水胁迫使植物体内氧自由基含量上升, SOD 酶活性也跟着上升。对照组 SOD 酶活性显著高于盐害组。对照组 SOD 酶活性一直在下降,提示盐害对其耐淹能力有很多的影响。
表 4 :淹水过程中叶肉细胞中超氧物歧化酶( SOD )活性的变化
淹水时间 对照组 盐害 1 盐害 2 盐害 3 盐害平均值
0 2.73 2.712 2.718 2.72 2.738
1 2.724 2.718 2.698 2.704 2.706
2 2.729 2.708 2.67 2.696 2.691
3 2.754 2.674 2.666 2.656 2.665
图 6 淹水过程中叶肉细胞中超氧物歧化酶( SOD )活性的变化 (A 曲线图, B 方柱图 )
A 图曲线 1 为对照组, 5 为盐害组的平均值变化。 B 图蓝色为对照组,褐色为盐害组平均值。
3 讨论
关于淹水胁迫下植物生理生化指标的变化,前人已经做过很多的研究。大致认为在淹水的过程中 MDA 的含量急剧上升; 涝害能使叶绿素的下降,同时可溶性蛋白质含量、 soD 和 cAT 活性均下降; M DA 含量和 PoD 活性上升。盐害造成植物因外界水势高而无法吸收,表现出与干旱相似 1 。我们的实验的出以下结论:
( 1 )从种子发芽后的 13 天的盐害胁迫使幼苗以后在外表上表现出受旱的特定。盐害组水稻的单颗重量要轻;叶片颜色也没有对照组绿。
( 2 )淹水胁迫下,水稻叶肉细胞形态模糊。叶肉细胞的液泡,叶绿体膜也逐渐模糊;这是由于缺氧和外界渗透压低于细胞,使得细胞膜吸水膨胀以致破裂。盐害组表现的尤其明显。这可能是由于在出苗期经过 13 天的盐害造成细胞膜结构不稳定和功能下降。 ( 3 )在 淹水的过程中,植物体内 MDA 和 POD 酶活性都会上升。其中盐害组 MDA 的含量显著高于对照组,上升趋势也更大。 植物器官衰老或在逆境下遭受伤害,往往发生膜脂过氧化作用,丙二醛 (MDA) 是膜脂过氧化的最终分解产物,其含量可以反映植物遭受逆境伤害的程度。 MDA 从膜上产生的位置释放出后,可以与蛋白质、核酸反应,从而丧失功能,还可使纤维素分子间的桥键松驰,或抑制蛋白质的合成。因此, MDA 的积累可能对膜和细胞造成一定的伤害。我们发现水淹对受过盐害的水稻的伤害明显大于对照的正常组。
( 4 )超氧物歧化酶( SOD )是植物体内清出自由基的重要酶。能防止膜被氧自由基破坏。本实验中盐害组 SOD 酶的活性明显低于对照组,其活性也一直下降。对照组 SOD 酶活性先下降后上升。盐害造成植物体内某些蛋白缺少,影响 SOD 酶的活性。植物体内存在一些能清除活性氧的酶系和抗氧化物质组分如 SOD 、 POD 等 , 他们构成了生物体的保护酶系统 , 并能协同作用抵抗外界环境胁迫。
从以上分析可以发现盐害对植物的抗淹能力有明显的抑制作用。其中可能是通过对早期幼苗生长的抑制作用来实现的。淹水逆境导致植物叶片膜脂过氧化作用增强 , 其主要氧化产物丙二醛含量增加 I 。这是由于淹水胁迫使植物体内产生大量的氧自由基 , 从而启动植物体内的膜脂过氧化作用 , 导致 MDA 含量增多。 MDA 的产生作为叶片衰老的标志 , 是衡量不同植物物种或品种耐淹水能力强弱的指标之一 , 本实验结果与此相符合。关于植物抗淹水机制的研究 , 有助于培育新的耐淹抗涝的作物 , 这对我国目前大力发展农业生产具有重要的实践意义。
参考文献:
? 张玉琼 , 张鹤英 锌营养对淹水玉米抗性的影响 安徽农学通报 1999,5(3):19-21 ? 魏和平,利容千,王建波 淹水对玉米叶片细胞超微结构的影响 植物学报 2000,42(8):811-817
? 倪君蒂,李振国 淹水对大豆生长的影响 大豆科学 2000 年 2 月 第 19 卷第 1 期
4 . 柴东方 裴凌鹏 李 伟 印莉萍 植物缺氧、缺铁胁迫的功能蛋白质组学 《生命的化
学》 2001 年 21 卷 5 期
5. 王金发,何炎明 《细胞生物学实验教程》 高等教育出版社 2002
Salty Inflence Damege to Rice by Flood
Qun JI Sheng-Tao Dai Tian-Hui Zhu Jin Shi Lei Zhang
( Life Science College of Sun-Yet Seng University , Guangzhou China ,510275)
Abstract:Varition of activation Catalase(CAT),SOD and MDA content under flooding stress were studied in leaves of rice.The results show that the MDA and POD both increase during flooding.
The cell of rice were destructed when flooded.
Keyword: damage of salty flooding stress sharp physylogy
范文五:珙桐叶肉细胞中的核内含体
() 植 物 学 报 1996, 38 6: 495, 496 A c ta B o ta n ica S in ica
研究简报
珙桐叶肉细胞中的核内含体
黄金生姜力甘习华朱伟华( ) 南京林业大学电镜室, 南京 210037
α 关键词 珙桐; 核内含体; 超微结构
INTRA NUCL EA R INCL US IO NS IN L EA F M ESO PHYLL
CELL S O F D AV ID IA IN VOL U CRA TA
2ΚΚ22H u an g J in sh en g J an g L iGan X ih u a an d Zh u W e ih u a
; ΨΨΓ210037E lectron M icroscop e L abora tory N anj ing F orestry U n iv ersity N an jing
A bstra c t T h e u lt ra st ru c tu re o f in t ran u c lea r in c lu sio n s in leaf m e sop h y ll ce lls o f D a 2
. v id ia in v olu c ra ta w a s in ve st iga ted w ith e lec t ro n m ic ro scop yIn t ran u c lea r in c lu sio n s o c2
. cu r gen e ra lly in th e ce lls o f yo u n g an d m a tu re leave sT h ey co n sist o f n um e ro u s b u n d le s
; ΓΚ10 agg rega ted b y seve ra l f ib re s d iam e te r abo u t nm som e t im e s a few o f b u n d le s tu rn to
. ; Γ2tu b u le s en ve lop ed b y f ib re sA u tho r s su gge sted th a t it is a n ew su b typ e F o f in t ran u2
.c lea r f ib r illa r in c lu sio n
ΜΜ Key word s D a v id ia in v olu c ra ta In t ran u c lea r in c lu sio n U lt ra st ru c tu re
1 对植物细胞核内含体的超微结构已有过广泛的研究。仅对玄参科就研究了 290 种植物, 发 B igazzi 2 现其中 242 种具有核内含体。根据超微结构特征, 可以把这类无膜的已证明主要是蛋白质成分的核内
(((()))) 含体分为 5 种类型。 即片状型 2、纤丝状型 2、管状型 2、晶体型 2及无定形L typ eF typ eT typ eC typ e 3 () 型 2。 此前除 2外陆续发现了与基本类型略有差异的 4 种亚型, 即 亚型、亚型、2 2 2A typ eF typ e C T A 2, 3 亚型和 亚型。 尽管目前还不清楚核内含体的功能, 但是研究发现, 它在成熟细胞中的结构是稳定 2 L 2, 4 的。许多学者认为核内含体作为超微结构特征, 在系统学和分类学研究中有重要价值。黄金生等对梓
5, 6 树属 3 种植物进行过研究。发现它们核内含体存在的情况一致, 也证明核内含体具有分类学意义。最
近我们在珙桐叶肉细胞中也发现了核内含体的存在。由于珙桐属在分科方面有一定争议, 因而这一发现
对探讨其分类位置及亲缘关系很有意义。
1 材 料 和 方 法
() 珙桐 叶片采自南京药物园。六月初采摘幼嫩叶片, 十月初采摘成熟叶片。取叶 D av id ia inv olu c ra ta 2 片中部在固定液中切成 1 左右小片, 投入有固定液的样品瓶中, 经轻微抽气使叶片沉入固定液中。 mm
() 使用 4% 的戊二醛和 1% 的四氧化锇 均用中性磷酸缓冲液配制双固定, 经乙醇系列脱水和环氧丙烷过
渡后, 用 环氧树脂渗透和包埋。 超薄切片厚度约 70 , 用醋酸铀和柠檬酸铅双染色, 最后在812 Epo n nm
2600 型电镜下观察与拍照。H
2 观 察 和 讨 论
小, 成熟细胞中内含体经常占据核的大部分位置。珙桐核内含体的轮廓没有一定形状, 也没有界膜, 在低倍率下似一缕缕的细丝。 在图版l|I 中, 由图 2 可以看到一缕缕的内含体交错排列, 在同一个核的切面上 可以同时见到细丝的纵切和横切。在较高的倍率下观察了核内含体的结构。图 3 显示出内含体的纵切面 是大量的纤丝, 纤丝有大的弯曲。 图 4 是内含体的横切面, 在横切面上清楚地看到纤丝并不是单根均匀 地分布, 而是聚集成束, 低倍下的细丝实际上是纤丝束。图 5 和图 6 是横切面的更高倍率相。图 5 是年轻
( ) 细胞中的核内含体, 可以测量出纤丝的直径约 10 , 若干根纤丝 大约 6, 9 根聚成一束, 偶尔可见纤 nm
丝束中纤丝紧密排列在束的外周, 呈现为中空的管状。 图 6 是成熟细胞中的核内含体, 与图 5 相比纤丝 束的排列混乱, 有更多的纤丝束变成管状, 此时大多无法分辨出组成管的纤丝。 以上观察表明珙桐叶肉 细胞在分化后迅速出现核内含体, 随着细胞的生长, 核内含体的体积变化不大, 只是组成内含体的纤丝 束有一定变化。
典型的纤丝状内含体在轮廓上是细长或梭形的, 由长而细小的波形纤丝组成, 横切面上直径为约 13 的纤丝单根均匀地分布。典型的管型内含体总是有规律的几何图形, 管的直径 22, 26 , 相互平 nm nm 行排列。珙桐中发现的内含体, 应属于纤丝状型, 但与典型的纤丝状型有明显的不同, 我们认为这是新发 现的纤丝状核内含体的亚型, 即 亚型。 亚型的特征是: 核内含体没有一定形状, 没有界膜, 由大量 2 2 F F
直径约 10 的纤丝形成细束或细管组成, 纤丝束或管有大的弯曲。nm
珙桐核内含体的发现有助于确定其分类学的位置。 为此我们将进一步对与珙桐属亲缘关系相近的 植物进行研究。
参 考 文 献
1 W e rg in W P ΚGube r P J ΚN ew com b E H. F ine st ruc tu ra l inve st iga t io n o f nuc lea r inc lu sio n in p lan t s. J U l tra R esΚ
1970. 30Π533, 577
2 . . B igazzi M A su rvey o n th e in t ranuc lea r inc lu sio n s in th e Sc rop h u la r iaceae and th e ir sy stem a t ic sign if icanceN ord J Κ1993. 13Π19, 31B ot . Κ. 3 B igazzi M T h e o ccu r rence o f in t ranuc lea r inc lu sio n s in th e L ab ia taeV e rbenaceae and Sc rop h u la r iaceaeC a ry olo2
g ia Ψ1984. 37Π269, 2924 Κ;Γ. Κ1982. 54Π. E sau K T ho r sch JN uc lea r c ry sta llo id s in sieve e lem en t s o f sp ec ie s o f E ch ium Bo rag inaceaeJ C el l S ci 149, 160
5 黄金生Κ樊汝汶Κ姜力Λ 梓树属植物花及叶细胞中核内含体的研究Λ 植物学报Κ1989Λ 31Π323, 3246 黄金生Κ姜力Κ甘习华Λ 黄金树核内包涵体的研究Λ 电子显微学报Κ1995Λ 14Π95, 98
图 版 说 明
( ) 图 版l|I 1、6. 成熟叶片的叶肉细胞。 2, 5. 幼嫩叶片的叶肉细胞。 11 两个相近的细胞中核内都有内含体 箭号。 ×4000 21 叶肉细胞的一部分。 在同一核的剖面上可以同时看到内含体的纵切和横切。 ×20000 31 核内含体的纵 切。×30000 41 核内含体的横切。内含体由大量纤丝聚成的细束所组成。×30000 5、61 核内含体横切的高倍率相。
1 年轻细胞中的核内含体。可分辨出纤丝的直径约 10 , 每 6, 9 根纤丝聚成一束, 有时个别的纤丝束 5×100000nm ( ) ( 呈现为管状 箭号。 61 成熟细胞中的核内含体。 纤丝束排列混乱, 出现更多的管, 组成管的纤丝大多不能辨认出 箭 ) 号。
E xp lan a t io n o f P la te
l|I . . 1, 6. . . 2, 5. P la te M e sop h y ll ce lls o f D av id ia inv olu cra taF ig sC e lls o f th e m a tu re leafF ig sC e lls o f th e yo ung
() . . 1. . ×4000 . 2. .leafF igIn t ranuc lea r inc lu sio n s a r row sin nuc le i o f tw o m e sop h y ll ce llsF igN uc leu s w ith inc lu sio n s
. ×20000 . 3. L eng th sec t io n and t ran sec t io n o f inc lu sio n s can be o b se rved o n th e sam e sec t io nF igL eng th sec t io n o f
th e inc lu sio n. ×30000 F ig. 4. T ran sec t io n o f th e inc lu sio n, show ing it co n sist s o f num e ro u s f ib re bund le s. ×30000
F ig s. 5, 6. H igh m agn if ica t io n o f c ro ss2sec t io n inc lu sio t s. ×100000 F ig. 5. In t ranuc lea r inc lu sio n in yo ung ce lls, show 2
( ) , 9 10 , 6ing th e bund le s a re agg rega ted by abo u t f ib re s d iam e te r abo u t nm som e t im e s a few o f bund le s tu rn to
(). . 6. , ,tubu le s a r row sF igIn t ranuc lea r inc lu sio n in m a tu re ce llsshow ing th e a r rangem en t o f f ib re bund le s is ir regu la r
( ) .m o re tubu le s app ea re and th e f ib re o n som e tubu le s a r row scan no t be d ist ingu ish ed ind ivedua lly
图版l|I 黄金生等: 珙桐叶肉细胞中的核内含体
2. : H u an g J in sh en g e t a lIn t ran u c lea r In c lu sio n s in L eaf M e sop h y ll C e lls o f
P la te l|I D a v id ia in v olu c ra ta
See exp lan a t io n a t th e en d o f tex t
转载请注明出处范文大全网 » 下图1表示绿色植物叶肉细胞部

