


 丿樱花灬
丿樱花灬
范文一:氨酰tRNA合成酶研究进展
20生物技术世界 BIOTECHWORLD
氨酰tRNA合成酶(ARS)作为生物进化初期出现的功能蛋白质 之一,不仅可以准确识别tRNA并使其氨酰化,还是生物进化中心法 则的基本保障。 根据结构特点不同,氨酰tRNA合成酶可分为Ⅰ、 Ⅱ 两类 [1]。 进一步细化其亲缘关系,每一类还可以细分到a、 b、 c三个亚 类。 其中,Ⅰ类合成酶因其独特的 “KMSKS” 和 “HIGH” 两个序列进 行Rossmann折叠,所以多以二聚体或是单体形式构成。 Ⅱ类酶具有 3段序列,第一段中的一个同二聚体构成了保守的脯氨酸残基。 在第 二段、 第三段中有精氨酸残基,脯氨酸残基、 精氨酸残基形成的活性 中心直接参与tRNA的结合及氨酰AMP的合成。
1 氨酰 -tRNA 合成酶种类
研究发现,AARS对蛋白质的合成意义重大,AARS功能不局限
于蛋白质翻译上,在生物进化过程中,扮演着极其重要的角色[2]。 生 物信息学以及结构生物学研究证实,AARS相互之间或同其他蛋白 质构成及其有序的一些列复合物,并参与生物体重要的生命生理过 程。
在古细菌、 细菌及真核生物中都发现了AARS复合物的存在。 古细菌中发现,AARS复合物包含很多种细胞因子及AARS,组成多 聚复合物,像亮氨酰-tRNA合成酶、 赖氨酰-tRNA合成酶、 脯氨酰-tRNA合成酶等等;而在细菌中,AARS复合物是由一个蛋白因子和 一个AARS组成的二聚体,脯氨酰-tRNA合成酶(ProRS)及YbaK复 合物能水解氨酰化的Cys-tRNAP ro;真核生物中,也发现了多种 含有AARS的多聚复合物,相比较而言,此类复合物比古细菌、 细菌 中含有的复合物更大更复杂,还具有广泛的生命功能。 其中已明确 了一些复合物的功能,像甲硫氨酰-tRNA合成酶、 谷氨酰tRNA-合 成酶同Arc1p蛋白N端偶联在生物体内,形成更加稳定复合物,利于 AARS催化和细胞核内tRNA向胞质运输;天冬氨酰-tRNA合成酶 和p38以及EF-1A的复合物可以提高氨酰化反应速度。
2 氨酰 -tRNA 合成酶特异性识别
为确保氨酰化反应高度专一性,AARS要从构成蛋白质的20种 氨基酸中及氨基酸类似物、 代谢物中专一识别某一种氨基酸,从众 多tRNA中识别同源tRNA。 tRNA分子大,可以提供多个和AARS特 异作用位点,因此,AARS容易对tRNA分子专一识别。 可是,对于很 小的氨基酸分子而言,再加上很多氨基酸侧链结构极为相似,又无 法提供充足的作用位点,所以AARS特异识别同源氨基酸较为困难。 学者已经发现一些AARS氨酰化中会活化一定频率的非同源氨基
酸,可是这些被活化的氨基酸再延伸到肽链中概率较低,所以这些 AARS形成编校结构域,可以水解误化氨基酸,进而保证遗传信息传 递的精确性。
3 氨酰 tRNA 合成酶的新功能
最新研究显示,氨酰tRNA合成酶不仅负责蛋白质合成,还参与 多种生命活动,包括DNA转录、 蛋白质翻译水平调控、 免疫应答和信 号传导等大方面 [3]。
在亮氨酸操纵子转录调控过程中,转录效率受到亮氨酰tRNA 合成酶对亮氨酸tRNA氨酰化水平的影响。 当亮氨酰tRNA合成酶的 活力缺失或表达较少后,大量未被酰化的亮氨酸tRNA集聚一起,促 进核酸在色氨酸操纵子翻译至14个氨基酸后发生停滞。 停滞诱导所 形成的一种抗终止高级结构可以阻碍核酸转录的结束,因此带动亮 氨酸操纵子增加表达。 然而,还没有氨酰化的亮氨酸能激活tRNAB. subtilis内的AT蛋白质,表达过量的AT蛋白能使TRAP结合亮氨酸 操纵子使其RNA活力丧失,所以亮氨酸操纵子的表达没有任何抑 制。 最近研究结果显示,亮氨酸tRNA调节是AT蛋白质是发生于转 录、 翻译双重生物学水平上的。
蛋白质合成效率同环境变化彼此适应就是蛋白质翻译调控的 效果。 原核生物中,很多RNA翻译合成蛋白质会受到反馈调节机制 影响。 如葡萄球菌中甲硫氨酰tRNA合成酶能抑制自身mRNA翻译, 此种负反馈抑制机制,生物学上称之为竞争性抑制。 甲硫氨酰tRNA 合成酶在mRNA的操纵子上链接,竞争地抑制结合核糖体30S亚基, 抑制mRNA翻译。 甲硫氨酰tRNA合成酶中mRNA操纵子共有4个结 构域,其中结构域1和3类似甲硫氨酸tRNA反密码茎环结构,能被甲 硫氨酰tRNA合成酶N端结构中的氨基酸识别。 在甲硫氨酰tRNA的 合成酶操纵子结构域2、 4中,核酸核糖体亚基就结合于2、 4部位中,当 甲硫氨酰tRNA的合成酶结合于结构域1、 3时,氨酰tRNA合成酶的 C端形成空间构象严重的阻碍核糖体亚基向结构域2、 4靠近。 上述实 验结果再次证实,甲硫氨酰tRNA合成酶合成生长速度具有依赖性 [4]。 生长速度增大,生物体内甲硫氨酸tRNA的浓度呈现正比例增长, 此时,甲硫氨酰tRNA合成酶发挥氨酰化功能,自身合成亦不再受到 任何抑制。
在真菌中,Neurospra crassa和其他细菌中线粒体色氨酰tRNA 合成酶为蛋白质Ⅰ类内含子剪切信号,具有类似作用的还包括酵母
氨酰 tRNA 合成酶研究进展
莫小 刘文丛 *
(吉林农业大学 吉林长春 130118)
摘要:氨酰tRNA合成酶作为生命进化中出现的最早的蛋白质之一,可促进氨基酸转移到tRNA上,参与生物体蛋白质合成。 伴随生物技术革命的 到来,氨酰tRNA合成酶内部结构以及生物功能成为国内外学者的研究热点。 在真核生物体内,氨酰tRNA合成酶以聚复合物形式表达功能,连接复杂的 生物分子体系。 氨酰tRNA合成酶作为蛋白质合成中最重要的一类酶,还直接参与、 调控、 RNA转录、 蛋白质翻译、 信号传导、 RNA剪接以及免疫应答等 许多生物的生命体征运动。 学者进一步还证实氨酰 tRNA 合成酶可以作为一种抗肿瘤药物潜在靶点。
关键词:氨酰tRNA合成酶 生物功能 蛋白质合成 中图分类号:Q816文献标识码:A 文章编号:1674-2060(2015)06-0020-01
作者简介:莫小(1989—),女,吉林长春人,硕士,研究方向:天然产物衍生化。
*通讯作者:刘文丛(1968—),男,吉林榆树,博士。
······下转第22页
范文二:氨酰_tRNA合成酶维持翻译忠实性的机制
ISSN 1007-7626 ttp: / /c jbmb. bjmu. edu. cnh中国生物化学与分子生物学报2011 年 2 月
CN 11-3870 /Q 27(2) :110 ,11 5Chinese Journal of Biocheistry and olecular Biology mM
??综述
tRNA -氨酰合成酶维持翻译忠实性的机制
* 谢兆辉
( ,253023)德州学院生物系山东 德州
-t . RNA摘要 氨酰合成酶在维持蛋白质合成忠实性方面具有重要的作用其忠实性机制可以分为
、、4 . t -RNA正确地选择底物转位前编辑顺式转位后编辑和反式转位后编辑 个水平不同的氨酰合
,tRNA ,-tRNA. 成酶能够利用其中的一种或几种机制将氨基酸和 连接起来形成正确的氨酰目, -tRNA ,,前氨酰合成酶的研究超出蛋白质合成已经延伸到了其它领域如致病机制和药物开发. -tRNA ,.等本 文就氨酰合成酶维持翻译忠实性的机制做一概述并对其应用前景进行了展望
;;;;关键词 蛋白质合成忠实性选择转位前编辑转位后编辑
Q75中图分类号5
Translational Fidelity Mechanism of Aminoacyl-tRNA Synthetases *XIE Zhao-Hui
( Department of Biology,Dezhou University,Dezhou 253023,China)
Abstract The translational fidelity can be influenced by factors at four levels: accurate selection ofsubstrate,pretransfer editing,cis-posttransfer editing and trans-posttransfer editing. Aminoacyl-tRNA synthetases ( aaRSs) play an important role in protein synthesis. Different aaRSs could follow one or more mechanism to load amino acids ( aa) to their accurate recognition tRNAs,and form aminoacyl-tRNAs ( aa-tRNA) . A t the present,the study of aaRSs has exceededb eyond their translational function,and extended to the fields of pathogenic mechanism and drug development. In this review,the translational fidelity mechanism of aaRS was discussed,and their application perspective of aaRS was speculated.
Key words protein synthesis; fidelity; selection; pretransfer editing; posttransfer editing
,遗传信息流动的每一步都具有高度的忠实性aaRS 1 的种类与进化. 所以差错率很低蛋白质合成的差错率是几个过程
aaRS,20 细胞质中一般有 种 它们分为两类,-tRNA 的净结果其 中起主要作用 的 是 氨 酰合 成 酶
,1,( ? ) ,. aaR( aminoacyl-synthetase, aaRS) .类与? 类各 自 起 源 于 一 个 祖 先由 于 ? 类和 核 糖 体S tRNA ,多催化小的或简单的氨基酸推测可能比 ? 类更古aaRS ,可以通过特异性的底物选择和编辑形成正,4, 确. aaRS aaRS,老原始 的结构小于现代 为单结构-tRNA( aa-tRNA) ,的氨酰核糖体则能够正确地选,aaS ,R域 蛋白类似于现代 的氨酰化结构域可以aa-tRNA . ,择 并合成肽链错误的氨基酸掺入肽链后识别
,核 糖体往往将肽链提前释放形成肽键合成后的一1 . aaRS 个 以 上 的 氨 基 酸现 代 都 含有附加的结构 ,2, ,5,. aaRS aa-种 忠实性 校 对 机 制而 错 误 负 载 的 . aaRS 域 结构域的 添 加 不 仅 增 加 了 第 二 种 忠 实,-tRNA ( AlaRS) 4 域如 丙 氨 酰合 成 酶 有 个 功 ,性机制而且还在调节氨酰化反应或转位前编辑中 t-RNA ,“可以通过密 码 子反 密 码 子 错 配以差 错 能 ,6, ”“”, 补 偿 或 差错协同的 方 式维 持 翻 译 的 忠 .-tRNA 具有重要作用 突变人胞浆亮氨酰合 成 酶 ,aaRS 实 性这 一 机 制 可能在进化上早于 的编辑机
收稿日期: 2010-10-27; 接受日期:2010-12-17,3,. aaRS ,制除 了 和 核 糖 体 之 外翻 译 的 延 伸 因 * Tel:13969214206; E-mail: xiezhh0523@ 163. 联系人
、tmRNA 子终 止 因 子 和 也对蛋白质合成的忠实com
Received: October 27,2010;Accepted: December 17,2010 . ,aaRS 性具有一定的作用目前的研究已经超出了* Corresponding author Tel: 13969214206; . aaRS 其在蛋白质翻译中的 作用本文主要就 维持E-mail: xiezhh0523@ 163. com
( LeuRS) ( CP1) . CysRS 编辑位点中保守的天冬氨酸利用锌离子与 了依赖锌离子的识别机制 Cys ( Asp) ,,7,,15,,. aaRSs ,Cys. ThrRS 不仅影响编辑活性也影响识别活性新结 的巯基作用专一性识别 大肠杆菌 的
,16,-,. ,-锌离子可以识别苏氨酸 γ羟基并排 除 缬 氨 酸构域的获 得 和 重 排 可 能 非 常 频 繁如 脯 氨 酰
Methanogenic ,( 不 同 于 其 它 生 物产 甲 烷 古 菌 tRNA ( ProRS) 合成酶家族获得编辑结构域的事 INS 2 ,生过 次因 为 其 编 辑 结 构 域 有 插 入 序 列
archaea) SerRS ( Ser) 的锌离子特异地与丝氨酸的氨 件至少发 ( insertion) 、N 末端附加和游离的编辑蛋白等多种形,17,8,,,Ser. SerRS 基结合从而特异识别 而酵母 则利用 . laS -t ( ThrS) ARRNAR式和苏氨酰合成酶的编
,18,,辑 结构域具有明显的序列相似性但插入酶中的tRNA Ser. SerRS 依赖性的方式识别 是 唯 一 进
,AlaRS ThrRS 位置 不同且 可能早于 获得编辑结erS ThrS SRR和 都利用锌离子识别氨基古菌的 aaRS,化 出两种机制识别同一种氨基酸的 因为产 ,酸,9,SerS ThrS RR所以推测 和 分化之前就已经存在甲烷 . ,构域这说明同一个编辑模块可以插入不同的酶,17, ..也可以 插入酶的不同位置 2 + ,-t ( spS) ,gRNAARM另外天冬氨酰合成酶中
aaRS 2 维持蛋白质翻译忠实性的机制Asp ,AspRS 通过 与 侧链作用影响 识别的专一性
,19,. aaRS :维持忠实性的机制大致分为两类特异性
. ,, 的底物选择和编辑氨基酸由于分子小结构相似,除氨酰化结构域能够特异性识别底物外编辑
,结构域也可以独立影响底物识别这在一定程度上 ,,-tRNA 往往难以区分而造成错误装载像异亮氨酰
aaS R说明现代 中的编辑结构域可能来源于独立存 IleS) ( R合成酶催化异亮氨酸装载 的 速 率 只 比 催 化
,在的编辑蛋白质这些独立编辑蛋白也需要特异性 ,10,200 . ,aaRS 缬氨酸高 倍为此很多 进化出了编辑 ,识别编辑底物这种识别特异性有一些仍保留在现 aaRS 的转位前编辑2. 2 . -AMP( aa-AMP) ,活性如果编辑发生在氨酰水平aaRS .代 中 aaRS ,转位前编辑也许是 选择反应的遗迹保存 ( Fig. 1 -) ;aa-称 为转位前编 辑 ?? 如 果 发 生 在 aaRS ,:aa-AMP 在了现代 中其方式主要有在氨酰
tRNA Fig. 1 ) . ,( 水平则称为转位后编辑 ? 现在已知化
( Fig. 1 ) 、aa-AMP 位点的选择性释放 ? 在 氨 酰 化 位 aaRS ,一半 左右的 具有编辑活性且大都同时具转位
t ( Fig. 1 ) 、aa-P RNAAM点非 依 赖 性 的 酶 解 ? 在 . 前和 转位后两种编辑活性转位前编辑多在氨酰化Fersht ,,域而 转 位 后 编 辑 只 在 编 辑 结 构 域所 以,11,结构 tRNA 、aa-AMP 氨 酰化位点 依赖性的酶解在编aaS .R“”提出了 编辑的双筛模型 等
2. 1 aaRS 对氨基酸的特异性选择 2. 2. 1 氨酰化中心的转位前编辑( Fig. 1) 转位前编辑的辑位点 的转位前编辑?和氨基酸的环化编
aaRS 特异性底物选择是 发挥忠实性功能的第 aa-P :AM第一种 方 式 为 的 选 择 性 释 放原 因 可 能 .辑等 是,aaRS,一步那些没有编辑活性的 则 只 能 靠 该 过 aa-AMP aaRS aa-非同 源 与 的 亲 和 力 比 同 源 . LeuRS 程 维 持 忠 实 性如 人 线 粒 体 和 啤 酒 酵 AMP . ProRS ,低容易从酶上释放到溶液中并水解( Saccharomyces cerevisiae ) -母 线粒体苯丙氨酰:tRNA 拥有两 种校对方式没 有 时可以酶 解或选tRNA ( PheRS) . aaRS 合 成酶对极性氨基酸的识,20,aa-P,. ,AM择性释放 但以选择性释放为 主另 外,“”,别往 往 运 用诱导契合机 制形 成 氨 基 酸 结
IleRS . 选 择 性 释 放也存在于 中转位前编辑第二种,合 口 袋从 而 氨 基酸残基与底物的极性功能团之
aa- AMP ,,. aaRS ,方式为 的 酶 解这 种 方 式 较 为 普 遍根 间形成氢键网络对非极性氨基酸的识别往
,往存在一个相对刚 性的结合口袋以增大与氨基酸tRNA :tRNA 据 是 否 依 赖 分为 两 类? 非 依 赖 性 ,12,,21,R-. ,aaRS 的 基范德华力作 用和堆积作用此外:ValRS ,的 酶 解这 最 早 在黄色羽扇豆的 中发现
,往往还进化出一些特 殊的识别机制以识别同源氨( Aquifex aeolicus ) LeuRS以后又在超嗜热 菌 的 ,. -tRNA 基酸或排除非同源氨 基酸酪氨酰合成酶,22,、大 肠 杆 菌 的 ( yrS) ,TR氨酰化中心具有 高度的亲水性能够特异,23, ,24,LeuRS ProRS . tRNA 和 中 发 现? 依 赖 性 的 酶 ,13,p-. ,性结合酪氨酸 羟基排除 苯丙氨酸真核和原核:LeuRS ,解细 菌 的 具 有 多 种 转 位 前 编 辑 活 性
23,,PheRS 生物 氨酰化结构域 均靠芳香族基团之间的tRNA . ,但 依 赖 性 的 酶 解 最 重 要此 外酵 母
,相互作用识别故只有芳香 族氨基酸才有可能成
112 第 27 卷中国生物化学与分子生物学报
( MetRS) ,DNA ,成酶使同 型 半胱氨 酰 环化成同型半胱聚合酶维持忠实性的一种机制这种机部分
氨 制,27,,28,. ,RNA aaRS .酸硫内酯除去另外焦磷酸解是 聚合酶和 也许对一些 的转位前编辑也有贡献
,29 , Fig. 1 The editing mechanism of aaRSPosttransfer editing; pretransfer editing in editing site? ?
( ES) ; Selective release of aa-AMP; aa-AMP hydrolysis in active site( ES)? ?
2. 2. 2 ThrRS ,编辑中心的转位前编辑同时存在 顺 式 和 反 式 两 种 转 位 后 编 辑形 成这种编辑形式最
aaRS “”. YbaK ProX 了 的三筛校对机制和 为 IleRS ,aa-中发现需将非同源 早在大肠杆菌的
AMPProRS
CP1,ValRS 从氨 酰 化 位 点 转 位 到 类 似 的 还 有 Pro Pro,33,,29,,,顺式编辑结 构 域 的 同 源 蛋 白游 离 存 在分 别 编 辑 LeuRCys- Ala-tRNA . YbaK ,. :CP1 和 存 在 于 细 菌 中证据有?的一些氨基酸残基突变会和 t RNAS aa-AMP ( Haemophilus influenzae ) ;的类似物结合在 最早在 流 感 嗜 血 杆 菌 中 发导致转位前 编 辑 丧 失? Pro ,转位后编辑结构域上转位前编辑和转位后编辑的 ,INS Ala-tRNA,YbaK 现可以 编 辑 则 通 过 反 式 ,23,Pro ,34,;IleRS ,位置重叠? 中可 能存在跨过氨酰化和 Cys-t,ProS . RNAR编 辑 形 成 的 第 三 个 筛 子
,al-P VAM编辑结构域的通道能 够 经 过 通 道 从 氨 酰 tRNA ,因 为 没有 结合特异性也不能同延伸因子,30, YbaK ProR通 过 和形 成 两元复合物发挥功 测 . 化位点扩散到达编辑位点但也有很多人对这种 ,竞争推 S ,35, . ProRS ProX 能的另一种反式编辑蛋白 主要在古:,CP1 方式存 有 疑 问首 先还 没 发 现 独 立 的 能 够 水
,ProRS 细菌和真核生物一些真核生物的 没有 aa-AMP. ,CP1 LeuRS 解 其次没有 的大肠杆菌 突 INS,,33,ProX ProRS . AlaRS 就形成了 第二个筛子是唯,6,,7,LeuS,R变 体和突变的人细胞质 仍然 可 以 进 一
. ,AN2690 LeuRS 行 转 位前编辑最近利用 封闭 编aaS,R广泛存在反式编辑蛋白的 所有生物的
,31,,32,,. Li 辑结构 域转位前编辑也仍然有活性但 等laS 1 1 . AR都有 个氨酰化结构域和 个编辑结构域另, ,认为也许后一种情况是第一种情况的一个特例因,laS AR外生 物界还 广 泛 存 在 的的反式编辑蛋aa- AMP 为 释放到溶 液 中 也 要 利 用 与 第 一 种 模 型 Ala AlaX ( AlaXp) ,ser-tRNAGly-白 反式 编 辑 和
,Li ValRS 一 样 的 通道且 等还提出了一种 转位前编Ala,5,tRNA. AlaXps Ser-所 有研 究 的 都 能 水 解
.辑的分 子机制 Ala Alat,laps lytRNAAXG-RNA只 有 少 数 可以水解 2. 3 aaRS 的转位后编辑 36,,. ,转位后编辑分为顺式编辑和反式编辑顺式编
ThrS,RN 大多数生物只 有 一 种 其 末 端 具 有 ,aaRS ,辑较为普遍发生在 的编辑结构域反式编辑
,比 较保守的编辑结构域而嗜热古菌和硫化叶菌. aaS R则发生在游离的编辑蛋白上大多数 只有一种 ThrRS,( ThrRS-cat) ,编码 两种 一种催化氨酰化
. ThrRS,反式编辑因为它具有反密码子结合结构域tRNA aaRS 3 在维持 翻译忠实性中的作用1 2 是目前发现的第 个进化出 种不同编辑结构域的
t aaS . ,RNAR首先可以影响 的底物选择性酵aaRS,ThrRS 因为古细菌的 编辑结构域与细菌和
,37,tRNA ,母 的结合可以改变氨酰化位点构象使 ThrRS .真 核生物 的不同源
SerRS ,具 有更高的专一性和催化效率并增加其对2 . 4 aaRS 转位前和转位后编辑的平衡
,18,. -tRNA 同源氨基 酸的特异 性大 豆 精 氨 酰合 成 aaRS 很多 同时具有转位前和转位后两种编辑
,活性但这两种形式在同一类酶中占的比重可以发 ( ArgRS) tRNA ,酶 结合刀豆的 时错 误装载刀 豆
. ,生变化那么两种编辑之间的平衡如何建立和维持 ,rgS t ARRNA氨酸的几率很 高但刀豆 和自身的 ,40, ,t RNA.区分刀豆 氨 酸 其 次能够影响转位前编 ,? aa-tRNA ,结合却能够有效 呢在核糖体校对 的 过 程 中校 对 时 间 越 . LeuRS ,tRNA 3′ CP1 辑中的 末端结合到酶的 结
. aaRS ,,23,长忠实性越高由于 催化的反应分为两步这 构 . 域上 是 转 移 前 编 辑 的 必 要 条 件 推 测 可 能 是
使人很容易想到这两步分割的时间会影响哪种编辑 t A76 RNA,的 核 糖 的 出 现增 加 了 活 性 位 点 溶 剂
,38,. 占主导现在很多实验都证明了转位前和转位后编 的 . ,有序性能够 更 好 地 稳 定 参 与 反 应 的 水 分 子 也
辑在 平 衡 中 占 的 比 重 主 要 决 定 于氨基酸转位给 tRNA ,有可能是 能 够 增 加 水 解 速 率或 作 为 自 发
,6,tRNA ,,的速率转位速率越快留给转位前编辑的时 水 AMP . aaRS ,,解的前奏加速了 的解离 在一些 中
,,间越短其在编辑中占的比重越小而转位后编辑占 tRNA 2′-OH 3′-OH 末端腺嘌呤核苷酸的 或 还直 接,aaRS 的比重越大这可能对具有独立编辑结构域的 ,“参与转位后编辑反应形成一种底物辅助型催,38, ,41,”化. IleRS ,Val-AMP 都适合如在大肠杆菌 中的水解 . . 机制这种机 制 最 早 在 古 细 菌 中 发 现 最 近 也 在
,LeuS ,1 R个 水 分 子被中 发 现编 辑 反 应 中 Val ,,IleRS 和 转位的速率几乎相同所以中 两 种 编 Leu 的tRN 3′′- ,3OH末端的 激活转位后编辑反应成为. ValRS ,Thr-AMP 辑都具有重要的意义而在 中的 AtRNA ,aaRS 一种 的自我切割这意味着 的转位后200 ,,ValRS 转 位速率是水解速率的 倍所以主要 编,38,,42,. ThrRS 以转 位后编辑 为 主大 肠 杆 菌 的 研 究 也 / . 辑是通过核 酶 蛋 白 质 杂合催化完成 的底 物 辅
,aa-AMP ,aaS R得 到 了相似 的 结 论氨 酰 化 位 点 的 动 力 助型催化在原始的酶中 非 常 普 遍的 这 种
学 分 割 是转位前编辑和转位后编辑平衡的决定因/ “”底 物辅 助 型 的 核 酶 蛋 白 质 杂 合 催 化 机 ,38,. ,aaS-R素可 能在非同源氨基酸活化过程中底物,aaS RRNA 制与 是从早期 世界最早进化出的一
,,复合物的 刚性结构降低容许一个水分子进入而其,类酶的观点一 致同时也可以说明转位后编辑也许
,他情况水 分子是排出的这也许是转位前编辑的基. t ,RNA有古老的起源除了上述作用之外还 可 以 通 .础 aa-过 影 响 , 两种编辑的平衡决定于氨酰基的转位的速率GlnRS AsnRS 和 的其中一种或两分古细菌缺乏 t . RNA合成维持翻译的忠实性一般真核生物和部分 那么影响转 位 速 率 的 因 素 就 会 影响哪种编辑占主 ,种aa-tRNA aa-tRNA,相关 的合成是先形成错 误 的 20 aaRS,原核生物有完 整 的 种 而 大 部 分 细 菌 和 . tRNA 导短时间内 平 衡 改 变 也 许 会 受 细 胞 内 和 氨 再 tRNA 以依赖性的转氨基方式形 成 正 确 部 aa- ,43,. ,基酸水平的影响而在进化过程中平衡的改变则可 tRNA . Thermus thermophilus ) ,( 嗜 热 细 菌 中延 伸Asn Gln Asn-tRNAGln-tRNA,aaRS 因子不结合 或 合成过能是由于酶自身的结构变化对其它功能的适
,39,,程 中 的错误中间 物体 现 出 具 有 翻 译 忠 实 性 的 功 . aaRS 应或编辑 结 构 域 插 入 位 点 差 异 等两 种 编 ,44,. aa-tRNAs ,能另外延伸因子与同源 的结合能力,辑的平衡也与编辑的氨基酸有关转位前编辑可以
远大于 ,aa-tRNAs ,,优先编辑一些氨基酸而转位后编辑则优先另一些 与错误 的 结 合 能 力推 测 在 某 些 情 况 下
aa-tRNA 延伸因子可以通过对 的特异性选择维持蛋. LeuRS 氨基酸最近人类胞浆 的相关实验也证明了
,31,.白质合成的忠实性 . LeuRS 这一点是研究转位前和转位后编辑的关
,CP1 系的一个理想模型除了具有保守的 编辑结构 4 展望
,LeuRS ,域外不同的 在 编 辑 机 制 上 差 别 很 大也 许 aaRS ,过去 的研究主要限于蛋白质合成现在其aa-AMP 这种编辑平衡的改变就是由于突 变 改 变 了
114 第 27 卷中国生物化学与分子生物学报
,45,2631 ;,辑位点在蛋 白 质 结 构 和 功 能 研 究 方 面通 过 改
Francklyn C S. DNA polymerases and aminoacyl-tRNA ,12,aaS,,R变 可以在蛋白质中引入非 天 然 氨 基 酸以 研
synthetases: shared echaniss for ensuring the fidelity of gene mm,究蛋白质结构和功能也可以产生自然界中不存在 ,46,expression,J,. Biochemistry,2008,47 (45) :11695-11703 ;,、的新蛋白质在病原学方面发现癌症自我免疫 ,13,Safro M G,Moor N A. Codases: fifty years after,J,. Mol Biol ,47,aaRS ,病和神 经 性 疾 病 等 都 与 特 定 有 关且 对 ( osk) ,2009,43 (2) :230-M 14,,aaRS 的 免 疫 症 状 已 成 为 了 鉴 定 ? 型 糖 尿 病 新 标 242 48,,;,aaS Fishan ,nkilova ,oor ,et al. Structure at 2. 6 RmRAVMNA 记在药物开发方面也开始成为新的药物
,7,,49,resolution of phenylalanyl-tRNA synthetase complexed with ,. 靶点如开发治 疗 疟 疾 药 物 的 新 靶 点现 在 抗
phenylalanyl-adenylate in the presence of manganese,J,. Acta 15,,aaS ( Bactroban) AN2690 R类 药 物 百 多 邦 和 已 经 Crystallogr Biol Crystallogr,2001,57 ( Pt 11) :1534-1544 D ,49,,aaS R用 于临床推测 药 物 开 发 将 成 为 研 究 最 Zhang C M,Christian T,Newberry K J,et al. Zinc-mediated
,aaRS 大 的 亮点相 信 的 深 入 研 究将 具有非常 ,16,amino acid discrimination in cysteinyl-tRNA synthetase,J ,. J
.深远的 意义 Mol Biol,2003,327 (5) :911-917
致谢 本文在撰写和修改过程中得到了焦传珍和张 ock-Bregeon C,ees B,Torres-Larios ,et al. chieving DA RAA
,!秀玲教授的大力支持在此表示衷心感谢 error-free translation; the mechanism of proofreading of threonyl- ,17, eferences)( R参考文献tRNA synthetase at atomic resolution,J,. Mol Cell,2004,16
(3) :375-386 ,1 , Ling J,So B R,Yadavall iS S,et al. Resampling and editing of
Bilokapic S,Maier T,Ahel D,et al. Structure of the unusual ischarged t prior to translation elongation,J,. ol Cell, mRNAM ,18,
seryl-tRNA synthetase reveals a distinct zinc-dependent mode of 2009,33 (5) :654-660
,2 ,Zaher H S,Green R. Quality control by the ribosome followingsubstrate recognition,J,. J,2006,25 (11) :2498-2509 EMBO
peptide bond foration,J,. ature,2009,457 (7226) :161-166 mN ,19,Gruic-Sovulj I,Landeka I,So? ll D,et al. tRNA-dependent amino 2 ,3 ,that bound Mgcontributes to amino acid and aminoacyl Seligmann H. Do anticodons of misacylated tRNAs preferentially acid discrimination by yeast seryl-tRNA synthetase,J,. Eur J + adenylate binding specificity in aspartyl-tRNA synthetase through mismatch codons coding for the misloaded amino acid( ,J ,. Biochem,2002,269 (21) :5271-5279 long range electrostatic interactions,J,. J Biol Chem,2006,281 BMC Mol Biol,2010,11 :41 Thompson D,Simonson T. Molecular dynamics simulations ,,4 (33) : 23792-Hartman H. Speculations on the origin of the genetic code,J,. J show 23803 ,20,Mol Evol,1995,40 (5) : 541-544 ,5 ,Hati S,Ziervogel B,Sternjohn J,et al. Pre-transfer editing by class Guo M,Chong Y E,Shapiro R,et al. Paradox of II prolyl-t synthetase: role of ainoacylation active site in RNAmmistranslation of serine for alanine caused by AlaRS recognition selective release o f noncognate amino acids,J ,. J Biol Chem, dilea,J,. ature,2009,462 (7274) :808-812 mmN ,6 ,,21,2006,281 (38) :27862-27872 Boniecki T,Vu T,Betha ,et al. CP1-M M A KJakubowski H. Valyl-tRNA synthetase form yellow lupin seeds: dependent partitioning of pretransfer and posttransfer editing in leucyl-hydrolysis of the enzyme- bound noncognate aminoacyl adenylate tRNA synthetase,J,. Proc Natl Acad Sci U S A,2008,105 as a possible mechanism of increasing specificity of the aminoacyl- ,7 , Pang L,artinis S . paradig shif tfor the aino acidY MAA mm( 49 ) : editing mechanism of human cytoplasmic leucyl-tRNA synthetase tRNA synthetase,J,. Biochemistry,1980,19 ( 22 ) : ,22,19223- ,J,. Biocheistry,2009,48 (38) :8958-8964 editing by class I leucyl-t synthetase,J, . J Biol Che,mRNAm5071-5078 19228 2009,284 (6) :3418-,8 , Ahel I,Korencic D,Ibba M,et al. Trans-editing of mischarged Zhu B,Yao P,Tan M,et al. tRNA-independent 3424 ,23,pretransfer tRNAs,J,. Proc Natl Acad Sci U S A,2003,100 (26 ) :15422- Tan M,Zhu B,Zhou X L,et al. tRNA-dependent pre-transfer 15427 editing by prokaryotic leucyl- tRNA synthetase ,J ,. J Biol Elucidation of ,9 , Beebe K,Ribas De Pouplana L,Schimmel P.Chem,2010,285 (5) :3235-3244 tRNA-dependent editing by a class II t RNA synthetase and ,24,
significance for cell viability,J,. EMBO J,2003,22 ( 3 ) :668- Splan , Ignatov , usier-Forsyth . Transfer K EM EMKRNA 675 modulates the editing mechanism used by class II prolyl-tRNA
,10,,25,Fersht A R. Editing mechanisms in protein synthesis. Rejection synthetase,J,. J iol he,2008,283 (11) :7128-7134 BCm iocheistry,Bmof valine by the isoleucyl- tRNA synthetase,J,.Baldwin A N, Berg P. Transfer ribonucleic acid-induced 1977,16 (5) :1025-1030 hydrolysis of valyladenylate bound to isoleucyl ribonucleic acid ,11,Fersht A R,Dingwall C.,26,Evidence for the double-sieve editing synthetase,J,. J Biol Chem,1966,241 (4) :839-845 mechanism in protein synthesis. Steric exclusion of isoleucine by
Jakubowski . isacylation of tLys with noncognate aino HMRNAmvalyl-tRNA synthetases,J,. Biochemistry,1979,18 (12 ) :2627-
acids by lysyl-tRNA synthetase ,J ,. Biochemistry,1999,38
,27,Kim H Y,Ghosh G,Schulman L H,et al. The cis-editing domain,J,. J Mol Biol,2009,394 (2) :286-296 relationshipDulic M,Cvetesic N,Perona J J,et al. Partitioning of tRNA- ,38,between synthetic and editing functions of the active site of an Proc atl cad Sci S A,NAU aminoacyl-tRNA synthetase,J, .dependent editing between pre- and post-transfer pathways in 1993,90 (24) :11553-11557 class I ainoacyl-t synthetases,J,. J Biol Che,2010,285 mRNAm
Airas R K. Effect of inorganic pyrophosphate on the pretransfer ,28, (31) :23799-proofreading in the isoleucyl-tRNA synthetase from Escherichia coli ,39,23809
,J,. Eur J Biochem,1992,210 (2) :451-454 Cusack S,arechuk ,Tukalo . The 2 crystal structure of YmAMA ,29,ing Jeynolds Ibba . inoacylt synthesis L,RN,MAm-RNAleucylt synthetase and its coplex ith a leucyladenylate -RNAmw- ,40,and translational quality control,J,. Annu Rev Microbiol,2009,analogue,J,. J,2000,19 (10) :2351-2361 EMBO 63 : Igloi Schieferayr . ino acid discriination by arginylG L,mEAmm- Insights into editing from an ,30,Silvian L F,Wang J,Steitz T A.61-78 ile tRNA synthetases as revealed by an examination of natural ile-tRNA synthetase structure with tRNAand mupirocin,J,.,41,
Science,1999,285 (5430) :1074-specificity variants,J,. FEBS J,2009,276 (5) :1307-1318 1077 ,31,,42,Schimmel P, Ribas de Pouplana L. Transfer RNA: from Chen X,Ma J J,Tan M,et al. Modular pathways for editing inihelix to genetic code,J,. Cell,1995,81 (7) :983-986 mnon-cognate amino acids by human cytoplasmic leucyl-tRNA Hagiwara Y,Field M J,Nureki O,et al. Editing mechanism ,43,synthetase,J,. Nucleic Acids Res,2010 Aug 30 ,Epub ahead of aminoacyl-tRNA synthetases operates by a hybrid ribozyme / ,32,of print, protein catalys,tJ,. J Che Soc,2010,132 (8) :2751-2758 Am m ,44,Li L,Yu L,Huang Q. Molecular trigger for pre-transfer editing Banerjee R,Chen S,Dare K,et al. tRNAs: cellular simulation study,J,.J Mol Model,2010 May 30 ,Epub pathway in Valyl-tRNA synthetase: A molecular dynamics aheadof print, barcodes for amino acids,J,. FEBS Lett,2010,584 (2) :387-Pro ,33,Ruan B,So?l l D.The bacterial YbaK protein is a Cys-tRNA395 Cys and Cys-tRNAdeacylase,45,J Biol Chem,2005,280 Cathopoulis T J, Chuawong P, endrickson T L. Conserved H (27) :,J,.discrimination against misacylated tRNAs by two mesophilic 25887-25891 Aminoacyl-tRNA synthetase ,34,elongation factor Tu orthologs ,J ,. Biochemistry, 2008, 47 ausann C DIbba . Hm, Mcomplexes: molecular multitasking revealed,J,. FEMS (29) :7610-7616 Microbiol ,46,Rev,2008,32 (4) :705-Ling J, So? ll D . Severe oxidative stress induces protein ,35,721 mistranslation through impairment of an aminoacyl-tRNA ,47,An S, Musier-Forsyth K. Cys-tRNA ( Pro ) editing by synthetase editing site,J,. Proc Natl Acad Sci U S A,2010,107 Haemophilus influenzae YbaK via a novel synthetase. YbaK. (9) :4028-4033 t ternary coplex, J, . J Biol Che,2005,280 ( 41) : RNAmm ,36,,48,Liu C C,Schultz P . dding new cheistries to the genetic GAm their connections to disease ,J,. Proc Natl Acad Sci U S A,34465-code,J,. Annu Rev Biochem,2010,79 :413-444 34472 2008,105 (32) :11043-Park S G,Park H S,Jeong I K,et al. Autoantibodies against 11049 ,37,,49,Sokabe M,Okada A,Yao M,et al. Molecular basis of alanine Bhat tT K, Kapil C, Khan S, et al. A genomic glimpse of aminoacyl-tRNA synthetase: novel diagnostic marker for type 1 discrimination in editing site,J,. Proc Natl Acad Sci U S A, aminoacyl-tRNA synthetases in malaria parasite Plasmodium diabetes ellitus,J,. Bioarkers,2010,15 (4) :358-366 mm2005,102 (33) :11669-falciparum,J,. BMC Genomics,2009,10 :644 Park S G,Schimmel P,Kim S. Aminoacyl tRNA synthetases 11674 and Shimizu S, Juan E C, Sato Y, et al. Two
complementary enzymes for threonylation of tRNA in
crenarchaeota: crystal structure of eropyru pernix threonyl-t AmRNA
synthetase lacking a
范文三:氨酰tRNA合成酶与周围神经变性及再生的关系
氨酰 tRNA 合成酶与周围神经变性及再生的关系
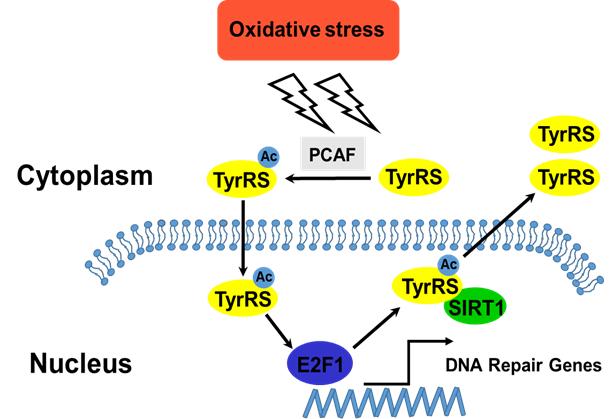
韩国庆熙大学医学院解剖学与神经生物学系 Junyang Jung教授所在研究小组初步结果表明氨 酰 -tRNA 合成酶对周围神经变性和雪旺细胞再生有影响。目前, Junyang Jung教授重点研究个别氨 酰 -tRNA 合成酶中在雪旺细胞神经变性和再生过程中的作用。但此研究目前还处于初级阶段,还 需要更多的研究来探索新的治疗周围脱髓鞘疾病和退行性神经疾病的方法。未来的研究应着眼于 确定哪些特定的氨酰 -tRNA 合成酶在周围神经变性和再生中起着必不可少的作用。 Jung 教授关于 这些研究的最新进展发表在《中国神经再生研究(英文版)》杂志 2015年第 8期文章中。
Article:
Jung J (2015) Aminoacyl tRNA synthetases and their relationships with peripheral nerve degeneration and regeneration. Neural Regen Res 10(8):1237-1238.
欲获更多资讯:请与《中国神经再生研究(英文版)》杂志国际发展部联络;联络电话:+8613804998773,或用电子 邮件联络:eic@nrren.org。 文章全文请见:http://www.nrronline.org/
氨酰 tRNA 合成酶与周围神经变性及再生的关系
作者所在研究小组初步结果表明氨酰 -tRNA 合成酶对周围神经变性和雪旺氏细胞再生的影响。目 前,作者的研究主要集中于神经变性和再生过程中调查个别氨酰 -tRNA 合成酶在雪旺氏细胞中的 作用。然而,如果开发新的治疗周围脱髓鞘疾病和退行性神经疾病策略 , 需要进行更多的实验研究。 这些研究应着眼于确定哪些特定的氨酰 -tRNA 合成酶是周围神经变性和再生的必不可少的要素。
英文摘要
Following damage resulting from mechanical injury, viral infection, or autoimmunity, peripheral nerves degenerate and a variety of complications, including sensory loss, muscular paralysis, skin thinning, and a loss of tendon reflexes, can manifest. If these complications persist, they can cause a number of debilitating personal and/or social problems. For example, Guillain-Barre syndrome is induced by the degradation of myelin sheaths, and if the symptoms are not controlled the disease can be life-threatening due to the demyelination of respiratory muscle nerves. To date, effective treatment strategies that can inhibit the demyelination of peripheral nerves in the early stages of
disease have yet to be developed. Thus, as with severe cases of Guillain-Barre syndrome, devices that alleviate the symptoms of respiratory muscular paralysis, including breathing machines, must be utilized until a cure can be developed.
范文四:ATP合成酶
ATP或称F0F1复合物(F0F1complexes),?该酶在分离状态下具有ATP水解酶的活性,在结合状态下具有ATP合酶的活性,?属F型ATPase。除了线粒体中有ATP合酶外,植物叶绿体的类囊体和好氧细菌都有ATP合酶的同源物,ATP合酶的分子组成和主要特点是:????头部:头部即F1,?细菌和线粒体ATP合酶的F1都是水溶性的蛋白,结构相似,由5种多肽(α、β、γ、δ和ε)组成的九聚体(α3β3γδε),α亚基和β亚基构成一种球形的排列,头部含有三个催化ATP合成的位点,每个β亚基含有一个。
柄部∶由F1的γ亚基和ε亚基构成柄部,将头部与基部连接起来。γ亚基穿过头部作为头部旋转的轴。构成基部的亚基b向外延伸成为柄部的构成部分。
基部∶基部称为F0,是由镶嵌在线粒体内膜的疏水性蛋白质所组成,由3种不同的亚基组成的十五聚体(1a:2b:12c)。其中c亚基在膜中形成物质运动的环,b亚基穿过柄部将F1固定;?a亚基是质子运输通道,允许质子跨膜运输。
范文五:蔗糖合成酶(SS)、蔗糖磷酸合成酶(SPS)
1.2.1 蔗糖合成酶(SS)、蔗糖磷酸合成酶(SPS)活性
参照於新建[]的方法并稍加改进。
酶液提取:取0.5g 左右预冷的水稻叶片(剪碎的去主叶脉的叶片) ,加4 mL缓冲液A(50mmol/L Tris—HC1,pH 7.0、10 mmol/L MgC12、2 mmol/L EDTA-Na2、20 mmol/L 巯基乙醇、2% 乙二醇) 于预冷的研钵中冰浴快速研磨成糊状,倒入离心管中,在低温冷冻离心机上4℃ 10000 r/rmin 离心30 min,所得上清液用于酶活性测定。
SS 活性的测定:在总体积0.15 mL的反应介质(含50 mmol/L Tris—HCL ,pH7.0、10 mmol/L MgC12, 10 mmol/L 果糖、3 mmol/L UDPG)中,加入100uL 酶液,30℃ 水浴中反应10 min ,加入2 mol/L NaOH 0.05mL,沸水煮10 min,流水冷却。再加入1.5 mL浓盐酸和0.5 mL 0.1% 的间苯二酚,摇匀后置于80℃ 水浴保温10 min,冷却后于480nm 处比色测定蔗糖的生成。活性单位以[蔗糖nmol /(g·min) ,Fw]表示。
SPS 活性的测定:在蔗糖合成酶反应体系中用10 mmol/L 果糖-6-磷酸取代10 mmol/L 果糖,其余均按蔗糖合成酶的方法。活性单位以[蔗糖umol /(g·h) ,Fw]表示。
药品配制(按100份)
酶液提取
1 配制缓冲液A---1000ml
200mmol /L Tris—HC1----500ml
200m mol/L Tris 12.1g----500ml 蒸馏水,
200m mol/L HC1/L 8.36ml 盐酸(37%)加水491.63ml 水
将 250 ml Tris+233.5 ml HC1,在此液中加入MgC12(分子量95.21,0.9521g ),EDTA-Na2(含两个结晶水分子量372,g ),巯基乙醇(分子量78,ml ),乙二醇(分子量,ml ),最后加蒸馏水定容到1000ml
2 配制缓冲液B---1000ml 5 ml
12.5ml Tris+11.65ml HCl,加入MgC12 0.0238g,果糖(259.19分子量,0.1296g ),加入UDPG (Uridine 5′-diphosphoglucose disodium salt from Saccharomyces cerevisiae分子量610.27,19.1ug ), 定容到25ml 容量瓶
12.5ml Tris+11.65ml HCl,加入MgC12 0.0238g,加入果糖-6-磷酸(304.1分子量,0.03041g ),UDPG (Uridine 5′-diphosphoglucose disodium salt from Saccharomyces cerevisiae分子量610.27, 19.1ug ), 定容到25ml 容量瓶
3 . 2 mol/L NaOH 50ml 0.4g 定容至50 ml
4. 0.1% 的间苯二酚 1g 999ml 蒸馏水
5.做蔗糖标准曲线
植物组织ATP 酶活性测定
一、原理
ATP 酶(adenosinetriphosphatase )可催化ATP 水解生成ADP 及无机磷的反应,这一反应放出大量能量,以供生物体进行各需能生命过程。它存在于生物细胞的多个部位,比如细胞质膜上,叶绿体类囊体膜上,对整个生命的维持有着重要的作用。在生物学研究中,常通过测定酶促反应释放的无机磷量或ATP 的减少量以及pH 变化等来测定ATP 酶的活力。本实验通过测酶促反应过程中无机磷的释放量来测定叶绿体偶联因子ATPase 的活力。偶联因子是分布在叶绿体类囊体膜表面的一种复合蛋白,它在光合作用能量转换反应中起重要作用。在正常情况下,膜上的偶联因子催化光合磷酸化反应(ATP 合成)的速率很高,而水解ATP 的活力是十分低的,但用二硫苏糖醇(DTT )、胰蛋白酶或较高温度等激活后,它水解ATP 的活力可大大增加。因此,偶联因子的测定常用激活后的ATPase 水解ATP 的活力来表示。
二、材料、仪器设备及试剂
(一)材料:新鲜菠菜10g ,洗净,去叶柄和中脉备用。
(二)仪器设备:1.分光光度计;2.水浴锅;3.照光设备(光源50,000Lx );4.台式离心机。
(三)试剂:1. Tris-HCl 缓冲液1mol/L(pH8.0):称60.57g Tris 溶于400ml 蒸馏水中,用浓盐酸调至pH8.0,再加蒸馏水至500ml 。2. 5mol/L 硫酸溶液:取27.8ml (比重1.
84)浓硫酸,慢慢加入到70ml 蒸馏水中,冷却后定容至100ml 。3. 10%硫酸钼酸铵溶液:称10g 钼酸铵溶于100ml5mol/L 硫酸中。4. 硫酸亚铁-钼酸铵试剂:称5g 硫酸亚铁,加入10ml 硫酸钼酸铵,再加蒸馏水稀释到70ml ,直至溶解为止(用前临时配制)。5. STN 缓冲液,将0.05mol/L Tris-HClpH7.8缓冲液(内含0.4mol/L 蔗糖、0.01mol/L NaCl) 放入冰箱中预冷。
三、实验步骤
(一)叶绿体制备及叶绿素含量测定:
取准备好的菠菜叶5g ,置于研钵或组织捣碎机杯中,加入20ml0℃下预冷的STN 缓冲液,很快研磨或捣碎(0.5min 完成),作成匀浆,以四层纱布过滤去粗渣,滤液于0~2℃下,200×g 离心约1min ,去细胞碎片,将上清液再于1500×g 离心5~7min ,取沉淀悬浮于少量STN (pH7.8)中,使叶绿素含量在0.5mg/ml 左右。小麦、水稻均可以此介质制叶绿体。如以甜菜叶子作材料,则匀浆介质中还应加入0.02mol/L 抗坏血酸钠。一般取0.1ml 叶绿体,加0.9ml 水和4ml 丙酮(分析纯),离心,取上清液于652nm 测吸光度,按Arnon 公式计算:叶绿素含量=A652×1000×5/(34.5×1000×0.1) =A652×1.45mg/ml
(二)ATP 酶的激活:
1. Mg2+-ATP酶激活液及反应液配制
2. 激活过程:取已制备好的叶绿体悬浮液1ml (叶绿素含量约为0.1mg/ml),加入1ml 激活液,于室温在白炽光50000Lx 下进行光激活6min 。
3. 反应过程:取三只试管,分别加入上述激活后的叶绿体悬浮液各0.5ml (剩下的叶绿体悬浮液供测定叶绿素含量用),再加入0.5ml 的反应液,取两管置37℃水浴中(另一管置冰浴中作空白用)保温10min ,各加入0.1ml20%的三氯乙酸停止反应。用台式离心机离心后各取上请液0.3~0.5ml (取样量按活力小大而改变)供测定ATP 水解的无机磷用。
转载请注明出处范文大全网 » 氨酰tRNA合成酶研究进展

