白日衣衫盡
白日衣衫盡
①蛋白质分子在行生物功能时,必须具有特定的二维空间结构。根据Anfinsen 理论,蛋白质分子的一级结决定其高级结构。蛋白质折叠问题就是研究蛋白质天然结构是如何形成的,即具有一氨基酸序列多肽如何逐步形成蛋白质所特有的空间结构。 ②蛋白折叠研究况:自从20世纪60年代,Anfinsen 基于还变性的牛胰RNase 在不需其他任何物质帮助下,仅通过去变性剂和还原剂就使其恢复天然结构的实验结果,提出了“多肽链的氨基酸序列含了形成其热力学上稳的天然构象所需的全部信息”的“自组装学说”。Anfinsen 的“自组装热力学说”得到了许多体外实验证明,的确有许多蛋白体外可行可逆的性和复,其是一些小分子量的蛋白,但是并非所有的蛋白都如。而且由于特殊的环境因,体内蛋白质的折远非。内蛋白质的折叠往往需要有其他辅助因子的参与,并伴随有ATP 水解。因此,Ellis 于1987年提出了蛋白折叠的“辅助性组装学说”。这表明蛋白质的折叠不仅仅是一热力学的过程,显然也受到动力的控制。科研人员已进行了大量出色的工作,但今为止我们对蛋白质的折叠机制的认识仍不完整的,至些方面还存在着错误的观。 ③研究蛋白质折的意义:⑴理论意义:蛋白质研究中最根本的科学问题就是多肽链的一级结构到如何决定它空结构?既然者决定后者,一级结构和空间结构之间肯定存在某种确定的关,有人把这设想的一级结构决定空间构的密叫作“第
码”。白质折叠机制的阐明揭示生命体内的第二套遗传密码,这是它的理论义。⑵实践意义:目前许多“象病”或称“折叠病”都是由蛋白质折叠异常造成分子聚集甚至沉淀引起的,如,老年呆症(蛋白质构象变化) 、囊性纤维病变(蛋白质不叠) 、家族性高胆固醇症(蛋白质错误折叠) 、家族性淀粉样蛋白症(蛋白质沉淀) 等等,因此深入理解蛋白质折叠机制这一需求已变得更加迫切。着蛋白质折叠研的深,人们会现更疾病的真正病因和更针对性的疗方法,设计更有效的药物。这是白质折叠研究的实
④蛋白质叠重要的潜在应用前景:⑴包涵体复性——利用DNA 重组技术可以将外源基因入主胞。但重组基因的表达产往往形成无活性的、不溶的包涵。折叠机制的阐明包涵体的复性会有重要帮助。⑵人设计蛋白——DNA 重组和多肽合技术的发展使我们能按照自己的计较长多肽链。但由于我们无法了解这一多肽将折叠为何种构象,从而无法按照自己意愿设计我们需要的、具有特定功能的蛋白质。⑶寻找致病机理——许多病,如阿兹海默症(Alzheimer's)、疯牛病(Mad Cow, BSE) 、可传播性海绵脑病(CJD)、肌萎缩性脊髓侧索硬化(ALS)、还有
(Parkinson's)等正是由于一些细胞内的重要蛋白发生突,导致蛋白聚沉或错误折叠而造成的。因此,入了蛋白质折叠与错误折叠的关系对些疾病的致病机制的阐明以及治疗方法的寻找将大有帮助。⑷揭示蛋白质功能——基因序列的发使我们得到了大量的白序列,结构信息的得对于揭示它们的生学功能是
要的。靠现有手段(X-ray 晶体衍射、NMR 及电镜)测定蛋白质的结构要较长的时间,因此结构解析的步伐已落后于发现新蛋白步伐。而结构预测的方法虽然速度较快,但可性不高,只有当我们对于维持蛋白质结构,驱动蛋白质折叠的理化因素更为了解,这一方法才可能有根本改进。另外,对于蛋白质相互作用、配与白质的作用等结构与能关系的研究也有赖于蛋质折叠机制
⑤关于分子伴:⑴分子伴侣可以定义为“一在序列没有相关性但有共同功能的白质,它们在细胞内帮助其他含多肽的结构完成正确的组装,而且在装毕后与之分离,不构成这蛋白质结构执行能时的组份”,如热休克蛋白。还有一“分子内伴侣”前肽常位于信号肽与成熟多肽间,是成熟多肽正确折叠所必需的。折叠酶则是催化蛋质折叠过程共键的异构化,主要有有PDI 和PPI 。的辅助叠分子可以是折叠酶又可是分子伴侣。⑵子伴在蛋质正叠中都起着关键性作用。首先,它可调节性地阻碍多肽链集聚;其次, 分侣还具有折叠互补功能,即可使未完全形成蛋白聚体的蛋白质重新折叠和复其水溶;此外,分子伴侣可以促进折叠错误的蛋白质降解。发现分子伴侣主是在蛋白质折叠的慢过程中发挥用。⑶分子伴侣本身不包括制正确折叠需的构象信息,而只阻止非天态多肽链内部的或相互间的非正确相互作用,或者说它们处于折叠中间态的多肽链提供了更多的正确折叠的机会,因而它提高折叠反应的产率而一定提高其
蛋白质折叠
蛋白质折叠
蛋白质折是生物化学和分子生物学的沿课题之一,近年来蛋白质折叠的研究日益引起人们注意的原是多的。其一,遗传信息由DNA 到RNA 再到蛋白质的程是分子物学的核心,通常称分子生物学的中心法则,经过多年的究人们对DNA 到RNA 再到多肽链过程已基本楚,但是白质的功能仅赖其一结构而且与空间结构紧密相关;其二,虽然蛋白质中一定的氨基酸顺序决定了其特定的空间结构的假说已被人们广泛接,但是怎样由一定的氨基酸排列的多肽链生成有一定的空间结构的白质的问题仍未解决。只有透地了解了多链是如通过自身内在的信息及与周围环境(包与各种蛋白质因子)的相互作用才能最终了解白质的空间结构与功
基因工程和蛋白质程是近年来生物技术发展的产物和先导,但们发现通基因工程和蛋白质工程所获得多肽有时并不能自身卷曲成有一定结构和完整生物学功能的蛋白质,其原因在于在多肽链的折叠上出了问题。此从基因程和蛋白质工程产翻译后加工的角也要求人们了解蛋
一、 蛋白质
关于氨基酸序列与白质空间结构的关系研究最早的工作是由C.Anfinsen (1960)关于核糖核酸酶研工作。他研究了核糖核酸酶的去和重折叠过程。该酶是由124 个氨基酸组成的蛋白质,有四对二硫键,其组合105{[(2×4)!/24×4!]=105}种可能方式。当用还原
(HOCH2-CH2-SH)时,二硫键被部分还原。继续加大b-巯基
硫键可部被还原。用8 M 的脲加b-巯基乙醇处理多肽链,分子内四对二硫键可全部被还原,链伸展为无规卷曲,酶活性完全丧失。但如果将脲和b-巯基乙醇透掉并在气中进行氧化,多肽链可又重新折叠为个具有三维构和催化活性的酶,它与未经处理的酶具有相同的溶解度并可结晶并获得相同的X-射线衍射图,其吸收光谱也相同。这是一个很好蛋白质一级结序决定其三结构例子,即顺序决定构象。Anfinsen 因此而获得1972年诺贝尔
二、关于蛋
各种实验理论计算均证明蛋白质的天构象在热力学上是最稳定的。那么一个具有特定的生物学活性和能的质究竟是如何找到这样一种热学稳定的构象的呢?这至今是一个未决的问题。我们可以一个由100 个氨基酸组成的小蛋白来进行讨论考虑:假设在这100 个氨酸组成的小白质中每个基酸残基有种同的象,那么总的构象数将是3100 =5×1047 ,如果从一种构象变为另一种构象所需要的时间为10 -13秒, 那么在上述的构象空间寻求一遍需要5×1047 ×10-13=5×1034 秒=1.6×1027 年! 而际上蛋质的折叠是在10-1~10 3 秒完成的。由此可见,蛋白质的折叠不是一个对种可能构象进行随机采
关于白质的折叠人
1. 框架模型(Framework model):
P. S. Kim 和R. L. Baldwin 于1982 年提出了蛋白质折叠的架模型,该模型认在蛋白质折叠的过程中大约有15个氨基酸基的多链首先折叠为瞬态的a 螺旋或b 层结构级结单元,然后这种瞬态的结构通过扩散彼此接近形成aa 、ab 或bb 的复合结构而获得稳定。这种复合构又称为折单元。折叠单作为一个核心引稳定其它摆动着的二级结单元,形成折叠框架,其它侧链将适应这
2. 疏
该模型提出折是由疏水折叠开始的,即四体石的疏水段首先聚集在一起,后一步聚集长大,形成一称为熔融球蛋白中间体。此种结构是一种具有二级结构但少有级相互作用的构,疏水残基
三、
蛋白质的去叠和重折叠亦即蛋白质变性复性。蛋白质的折态只有在最适条件下才可存在。环境的改变,诸如温度、pH 、变剂、压力等作用都可使
变性的物理础是:pH 的改变可使盐键裂,使埋藏在蛋白质结构内部的非解离基团得到裂解而暴露出来,蛋质去以减少静电相互作用;变性剂如脲盐酸胍等可使蛋白质结构中的键发生断,这增加了非极性分子括氨基酸侧链的溶解度,减少了疏水相互用;脲还可深入到蛋白质分子的内部影响蛋白分子的密堆积。此外,去污、有机溶剂、金、热、机、冷冻、超声、高压、辐射等均可引起蛋白质的变性。些性均不会破坏蛋白质的共价键而是只涉及到氢键、盐键、疏相互作、范得华相互作用等次级键的破坏。有些变性的蛋白当变性因素被除去后又自动地恢复到天然状态,这种现称为蛋白质的复(renaturation),这种复性即蛋白质叠研究中的重折叠(refolding)。(1931年吴宪提出的蛋白
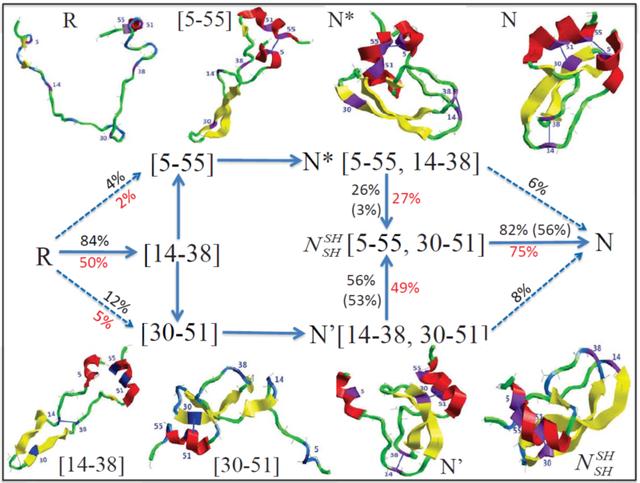
长期以来于蛋白质的去折叠与折流行着一种二态模型。即认为在蛋白质变性过程中次级键间着一种协同作用,当变性因增强时最初并不能观察蛋白质分的结构有何变化,当达到某一阈值时由于某些关键的次级键变化而导致蛋白质构象的急变化。二态模型可表示U 、N 相转化的, U 代表伸展态,N 代表天然态。二态模型认为从伸展态到天然态的过程或反之过程是一个完全协调的过程,没有可观察到的中间态。但近年Baldwin 等的研究发现,以上二态型需要进行正。他利用重氢交换快速混合技术以及二核磁波谱技术等有效地捕获了结构稳定的叠中间产物。具体
1 将白质溶于含4.2M GuHCl pD 值为6.0的重水中,此时蛋白质多肽链完全伸展所有
2 入0.1M 醋酸水
3 经时间tf ,入pH 值为9.3 的缓冲液,使所有溶剂可接近的质子在1ms 的时
4 降pH 值使交换止,再折叠过成。利用二维核磁共振技术,分辨哪些残基被氘交换并形成了
通过改变tf 可以确定哪些部分先于其它部分叠,还可示不同中间体形成的序。例如,利用此技术他们明了细胞色素C 折叠的第一步为多肽链链两端的两个螺旋形成,核糖核酸酶的叠则是分子中
四、蛋白
近年人们对蛋白质的热力
质是一个成千上万个原子组成的观体系,它既有序又乏称性。对它的了解象对其它热力学体系一样,仅了解它的构是远不够的,必须了
蛋白质构的稳定性可蛋白质的去折程的自由能来表示,如以N 表示天然构象,以D 表示
N 、D
天然态和伸展
ΔG= GD-GN
由各种计算方法,人们可以计算在天然态和变性态的热量和定热容、D>N 的数值、与溶剂的接触、有序等,可通过这些计算结果作系列的氢键断裂、折叠程度、温度与有序度。以及在某一温度下与溶剂接的影响曲线,从而研究某体的蛋白质的折稳定性及其与周围
近来对于解蛋白质是如何快速的形成能量低状即天然状态,一个被接受的模型是“漏斗模型”,即在一个漏斗状的能量表面上,蛋白可能通一系不同的路径行折叠。这
五、蛋
近年来已经报道了一些新方法成功地由氨基酸顺序信息预测了它们折叠构象。了提供蛋白质结构预测的标准,1994 年来自世界各地的35个实的约150 名科学家在美国加州的Asilomar 开会对33 个蛋白质结的预测问进行总结。这些要行构预测的蛋白质已知折叠和结构的蛋
蛋白,每个实验室进行诸如蛋白质的结构类型、二级结构和子坐标(三级构)等的预测。在分配给各实验室进行构预测时根据不同的情况向每个实验室提供诸如蛋白质名称、氨基酸顺序、晶体学家建议和该蛋的考文献等。利用这些信息加上蛋白质结的各种数库,有的验室的目标据同源蛋白的结构建立详细的三维结构模;有的实验室则只需行二级结构的预测等。所有这些预测的结至少在会议前的一个月提交。由会议的结果获得了有关蛋白质折叠的识别问题,即将一个新的顺序与一已知的特征化的维结构花样的叠合识别在大多数情况下是比较成的,在某情况下是相可靠的,尤其是当两个蛋具有相同的重要氨基酸顺时,它总是在那区域具同的三维结构。对于两个同源蛋白的折叠识别就如同出这两个蛋白一个未知蛋[Test Protein]一个已知蛋白[Template Protein] 的氨基酸最佳对齐一样简单。当两蛋白间的氨酸顺序相同性低时,不论氨基酸残基间的相似性如何,这些氨基酸残基仍据在等效的位置。例如在未知蛋中的一个异亮氨酸(Ile)取代已知蛋白中的氨酸(Val),这两个蛋白的折叠仍然相同的。然在白质三维结构的相同位上果一个精氨酸取缬氨酸会影响折叠,但如果缬氨酸暴露于蛋白质的分子表面的话,则这种取代也会对折叠有大影。实际上需要的只是估计出没有同源性的两个蛋白之间的顺序相似性以计已知蛋白中的每个位置上的可允的取代加上插入
六、蛋白
人们发折叠识别的方式之一通过比较,从进化分叉机理上紧密相关蛋白的族列出一个直接了当的取代/插入/缺失(sub-in-del)的表,然后进行对齐比较,这种对齐接表示在一致性顺序(consensus sequence) 个位置的功能容忍性与由进化所产生的顺序变化之间的关系。如果蛋白家族相对于氨基酸顺序的变化占主导地位,则基本可断出其所有成员结构特性,并测出折叠的共特征。此外从取代/插入/缺失(sub-in-del)表中的多重齐顺序可识别出
插入/缺片断(环和非结构连片断)和那些不能忍插入/缺失片断(a 螺旋和b 链),再加上可识的b 链a 螺旋和暴
基于这样一致性模型的结果加州学旧金山分校的Fred Cohen 举出了五种更同源蛋白的家族在没有晶结构的情况下根据二级构和环的测方式,推测出了它们的结构。例如,他们把丙酮磷酸二激(pyruvate phosphate dikinase )的第个结构域(domain 4) 正确地定位于具有a/b TIM 桶花样的蛋白类型中;另外有其它的几个组也正确地预测了Chorismate Mutase (变位酶)一个a 结构的蛋白,还有个组预测出该蛋白的两个相同亚基通过a-coiled-coil 方式作为二体间相
这种预测方法的最显的特点是利用了可由同源蛋白家族序列所推断的量的结构信。虽然对于二级结构的预测似乎有高准确性,但往往有可能发生的严重,即把a 螺旋预测为b 折叠,或反之。利用同源蛋白家族序列就使得这种发生严错误的可性大大降低;此外由环域常常具有一个或个插入/缺失因而可好地进行
另一种折的识别方式甚至是更成功的,即所谓的“Threading”。此方式主要是通过分析知蛋的实验所测得的三维结构来估可允许的sub -in-del ,非进行未知结构与同源家族的多重对齐。早期的threading 方式之一是把每个位置的局部境根据包埋水和二级结分布情况分几,过蛋白质结构数据以及在每种环境类型下20 种氨基酸残基中的每种残基占据的经验可能性,用作取代可能性的分值。近来这种研究已被另一种方法所取代,即用已蛋白质结构中的氨基类型的经验配对势的小来衡取代的可能。简言,一个位置上可取代的容忍度是通过某位置上和其残基周围的所有位置上的氨基酸类间的相互作用自由能
Threading 的过程包括将未知蛋白与具有最高分值的已知蛋相对齐。为了之与未知蛋白最好地吻合对齐,要阅知模板蛋白的文库。具有高分值的蛋白的折叠就可被考虑为最佳的未知蛋白的折叠的候选者。当用未知顺序本身来估计配势,以及未知顺序中的所有可间在模板上threading 时,仅是一计算时间
最激动心的基于threading 的预测是对一些已知蛋白(已知折叠方式)的预都获得了极高的功率。所预测的这些折叠形式包括4 个TIM 桶的结构的蛋白枯草杆菌蛋白酶(Subtilisn )的前肽;Synaptotagmin (一种类免疫球蛋白)折叠;rtp (复制终止蛋白)等。有一个究组从9 预测中获得了5 正确的预测结果。当然出现了不同的两个组获得的论出现矛盾
蛋白质折叠
蛋白质折叠
结构决定功能,仅仅知道基因组序列并不能使们充分解蛋白质的功能,更法道它是如何工作的。白质可凭借相互作用在细胞环境(特定的酸碱度、温度) 自己组装自己,这种自我组装
蛋白质折叠题被列为“21世纪的生物物理”的重要课题,它是分子生物学中心法则尚未解决的一个重大生物学问题。从一序预测蛋白质分子的三级结构并进一预测其功能,是极富挑战性的工。研究蛋白折叠,尤其是折叠早期程,即新生肽段的折叠过程是全面的最终阐明心法则的一个本问题,在这一领域中,近年来的新现对生肽段够自发进行折的传统概念了根的修。中,X 射线晶体衍射和各种波谱技术以及电子显微镜技等了极其重要的作用。第十三届国际生物物理大会上,Nobel 奖获得Ernst 在报告中强调指出,NMR 用于研究蛋质的一个主要优点在于它能为详细的研究蛋白质分的动力学,即动态的结构或构的运与蛋白质分子功能的关系。目前的NMR 技已经能够在秒到皮秒的时间域上观察蛋白质结构的运动,其中包括主链和侧链
在各种不的温度和压力下蛋白质的叠和去折叠过程。蛋白质大分子的结构分析也不仅仅只是解某个的结构,而是更加关注结构涨落和运动。例如,运输分子的酶蛋白质通常存在着种构象,结合配体的和未结合配体的。一种构象的结构涨落是构象转变所必需前奏,因需要把光谱,波谱学X 线构析结合起来研究结构涨落的平衡,构象改变和改变过程中形成的多种中间态,又如,为了了解蛋白质是如何折叠的,就必须知道折叠时几个基本过程的时间尺度机制,包括二级结构(螺旋和折叠)的形,卷曲,长程相互作以及未叠肽段的全面崩溃。多种技术用于研次过程,如快速核磁共振,快速光谱技术(光,远紫外和近紫外
【1蛋白
在生物体,生物信息的流动可以分两个部分:第一部分是存储于DNA 序列中的遗传信通过和翻译传入蛋白质的一级序中,这是一维信息之间传递,三子密码介导了这一递过程;第二部分是肽链经过疏水缩、空间曲、侧链聚集等折叠过程形蛋白质的天然构象,同获得生物性,从而将信息表达出来;而蛋白质作为生命信息的表达载体,它折叠所形成的特定空间结构是其具有生物学功能的基础,也就是说,这个一维信息向三维信息的化过程是表现生命活所必需的。 从20纪60年代,Anfinsen 基于还原变性的牛胰RNase 在不需其他任何物质帮助下,仅过去除变性剂和还
恢复天结构的实验结果,提出了“多肽链的氨基酸序列包含了形成其热力学上稳定的天然构所必需的全部信息”的“自组装学说”以来,随着对蛋白质折叠究的广开展,人们对蛋白质折叠理论有了进步的补扩展。Anfinsen 的“自组装热力学假说”得到了许多体外实验的证明,的确有许多蛋白在体外可进行可逆变性和复性,尤其是一些小子量的蛋白,是非所有的蛋白都如此。而由于特殊的环境因素,体内白质的折叠远
体内蛋白质折叠往往需要有其他辅助因的参与,并伴随有ATP 的水。因此,Ellis 于1987年提出了蛋白质折叠“辅助装学说”。这表明蛋白质的折叠不仅仅一个热力学的过程,显然也受到动力的控制。有学者基于有些相似氨基酸列的蛋白质具有不同的折叠结构,而另外一些不同基酸序列的蛋质在结构上却相似的现象,提出了mRNA 二结构可作为一种遗传密从而影响蛋白结构假说。但目止,该假说尚没有任何实验证据,只有一些纯数学论证[3]。么,白质的氨基酸序列究竟是如何确定其空间构的呢?围绕这一问题科人员已进了大量出色的工作,但迄今为止我们对蛋白质的折叠机制的识仍是不完整的,甚至有些方还存在着错误的观点。 在这方面出重要贡献的典研究实例美国C.B. 安芬森小组关于牛胰核糖核酸酶的变和复性的研究。牛胰核糖核酸酶含有124个氨基酸残基, 由8个巯基配对组成4对二
算出酶子中8个巯基组成4对二硫键的可能方式有105种, 这就提供了一个定量估算复性重组的指。在温和的碱性条下,8摩尔的浓脲和大量巯基乙醇能使四对二硫键全还原,整个分子变为无规则卷曲状,酶分子变性。透析去,在氧存在下,二硫键重新形成,酶分子完全复性,二硫键中成对的巯基都与天然一样,复性分子可以结晶且具有与天然酶晶相同的X 射线射花样,从而实,分子在复过中, 不仅能自发地重新折, 而且只选择了105种二硫可能配对方式中
【2蛋白质
▲框
框架模[4] 假设蛋质的局部构象依赖于局部的氨基酸序列。在多肽链折叠过程的起阶段, 先迅速成不稳定的二级结构单元; 称为“flickering cluster”, 随后这二级构近接, 从而形成稳定的二级结构框架;最后,二级结构框架相互拼接,肽链逐渐紧缩,形成了蛋白质的三级构。这个模型为即使是一小分子的蛋也可一部分一部分的进行折, 其间形成的亚结构域是折中间体的重
▲疏
在疏水塌模型[5]中,疏水作力被认为是在蛋白质叠程中起决定性作用的力的因素。在形成任何二级结构和三结构前首先发生很快的
▲扩散-碰撞-粘合机制 (Diffusion-Collision-Adhesion Model )
该模型为蛋白质的折叠起于伸展肽链上的几个位点,在这些位点上生成不稳定的二级结构单元者疏水簇,主要依局部序列的进程或中程(3-4个残基)相互作来维。它们以非特异性布朗运动的方式扩散、碰撞、互附,致大的结构生成并因此而增加了稳定性。进一步的碰撞形成具有疏水核心和二级结构的类熔球态中间体的球状结构。球形中间体调为致密的、活性的类似天然构高度有序熔球态结构。最后活性的高度有序熔球态转变为整的有活力的
▲成核-凝聚-生长模型(Nuclear-Condensation-Growth Model )
根据这种模型,肽中的某一区域可以形成“折叠晶核”,以它们为心,整个肽继续折叠进而获得天然构象。所“晶”实际上是由一些特殊的氨基酸形成的类似于天然态相互作用的网络结构,这些残基间不是以非特异的疏水作用维的,而是特异的相互作用使这残形成了紧密堆积。晶核的形成是折叠起阶段限速
▲拼
此模型[9]的中心思想就是多肽链可沿多条同的途径进行折叠, 在沿每条途径折叠的过程中都是天然结构越来越多, 最终都能形成然构, 而且沿每条途径的
一途折叠方式相比, 多肽链速度较快, 另一方面, 外界生理生化境的微小变或突变等因素可能会给单一折叠途径成大的影响,而对具有多条途径的折而言, 这些变化可能给某条折叠途径带来影响, 但不会影响另外的折叠途径, 因不会从总体干扰多肽链的折叠, 除这些因素造成的变太大以致于从根本上响多肽链的
【分子伴侣】
1978 年,Laskey 在进行组蛋和DNA 在体外生理离子强度实验发现, 必须要有一种细胞核内的酸性蛋白———核质素(nucleoplasmin) 存在时, 二者才能组成核小体, 否则就发生沉淀。据此Laskey 它为“分子伴侣”。分子伴侣指能够结合和稳定另外一种蛋白质的不稳定构象, 能通过有控制的合和释放, 促进新生多肽链的折叠、多聚的装或降解及胞器蛋白的跨膜输的一类蛋白质 [10,11] 。子伴侣是从功能上定义的, 凡具有这种功能的蛋白质都是分子伴侣, 它结构可以完全不同。这一概念目前已延伸到许白质, 现已鉴定出来的子伴侣主属于三类高度保守的蛋白质家族[12]:stress 90 family 、stress 70 family、stress 60 family。中stress 60 family存在于真核生物的线粒体(在乳动物中称为Hsp58)、叶绿体(称为cpn60)中,在生物的细胞质中,它
【意义和前景】
蛋白质折机制的阐明将揭示生体内的第二套遗传密码,这是它的理论意义。蛋白质叠研究,比较狭义的定义就是究蛋白质特定三维空间构形成规律、稳定性和与其生物活性的关系。在概念上有力学的题和动力学的问题;蛋白质体外折叠和在细胞内叠的问;理论研和实验研究的问题。这里最根本的科学问题就是多肽链的一级结构到底如何决定它的空间结构?既然前者决定后者,一级结构和空间结构之间定存在某种确定的系,这是否也像苷酸过“三联密”决氨基酸顺序那样有一套密码呢?人把这设想的一级结构决定空间结构的码叫作“第二遗
如果说“三密码”已被破译而实际上已成明码,那么破译“第二遗传密码”正是“蛋白质结构预测”从理论上直接解决蛋白质的折叠问题,这是蛋白研究最后几个尚未揭示的奥秘一。“蛋质结构预测”属于理论面的热力学问题。就是根据测得的蛋白质一级序列预由Anfinsen 原理决定的定的空间结。蛋白质氨基序列,特别编白的酸序列的测定现在几乎已经成为常规技术,从互补DNA (cDNA )序列可以根据“三联密码”推定氨基酸序列,这在上世纪获得重大突破的分子生物学技术,大大加速了蛋质一级结构的测定。目前白质数据库中已经存大约17个蛋白的一级构,但测定了空间结构的蛋白大约只有1.2万个,这中间有许多是很相似的同源蛋白,而真正不同的白只有1000多个。
组计划的胜完成,解读了人类DNA 的序列,蛋白质一级结构的数长必定会出现爆炸的态势,而空间结构测定的速度远远滞后,因此二者之还会成更大的距,这就更需
同时,它还存重要的潜在应用前景,例如以几个方面: ▲利用DNA 组技术可以将外源基因入宿主细胞。但重组基因的表达产物往往形成无活性的、溶解包涵体。折机制的阐明对
▲DNA重和多肽合成技术的发展使我能够照自己的意愿设计的多肽链。但由于我们无法了解这一多肽将折叠为何种构象,从而无按照己意愿设计我们需要的、
▲许多病,如阿兹海症(Alzheimer's),疯牛病(Mad Cow, BSE) ,可传播性海绵状脑病(CJD),肌萎性脊髓索硬化症(ALS),还有帕金氏症(Parkinson's)等正是由于一些细胞内的重要蛋白发生突变,导致蛋白质聚沉或错误折叠而造的。因此,入了解蛋白质折叠与错误叠关系对于这些疾病的病机制的阐明以及治疗方的寻找将大
▲基因组序列的展使我们得到了大量的蛋白质序列,结构信息获得对于揭示它们的生物功是十分重要的。依靠现手段(X-ray 晶体衍射、NMR 及电镜)测定蛋白质的结需要较的时间,因此构解析的步伐已后于发现新蛋
构预测的方法虽速度较快,但可靠性并不高,只有我们对于持蛋白质结构,驱动蛋白折的理化因素更为了解,这方法才可能有根本的改进。另外,我们对于蛋白质相互作用、配与蛋白的作用等结构与能关系的研究也赖于蛋白质折
【蛋白质折
人们对由于基因突造成蛋白质分子中仅仅一个氨基酸残基的变化就起疾病的情况有所了解,即所谓“分子病”,地海镰刀状红血球贫血症就是因为血白分子中第六位的谷氨酸突变成了颉氨酸。现在则发现蛋白质分子的氨基酸序列没有变,只是结构或者说构象有改也能引起疾病,那是所谓“构象病”,称“折叠
大家都道的疯牛病,它由一种称为Prion 的蛋白质的感染引起的,这种蛋白质也以感染人而引起神系统疾病。在正常机体中,Prion 是常神活动所需要的蛋白质,而致病Prion 与Prion 的一级结构完全相同,只是空间结构不同。这一疾病的研究涉及到许多生物学的基本问题。一级结构全相同的蛋质为什么会不同的空间结,这Anfinsen 原是否矛盾?显然这里有蛋白质能量和稳定
从来认为蛋结构的变化来自于序列的化,序列的变化来自于的变化,生命信息从核酸传递到蛋白。而致病Prion 的信息被诺尔奖获得者普鲁辛纳证
致病蛋白Prion 导致正常蛋白Prion 转变为致病的叠状态是通蛋白分子间的作用而感染!这相作用的本质和机制是什么?仅变了折叠状态的分子又如何导致严重的疾病?这些问题都不能用传统的概念给满意的解,因此在科学界引烈的争论,有关究的强度和竞争性
由于蛋白质折异常而造成分子聚集甚至沉淀或能正常转到位所引起的疾病还有老年痴呆症、囊性纤维病变、家族性高胆固醇症、家族性淀粉样蛋白、某些瘤、白内障等等。由于分子伴侣在白质折叠中至重要的作,分子伴侣本身的突变显然会引蛋白质折叠异而引起折叠病。随着蛋白质折叠究的深入,人们会发现更多疾病的真正病因和更针对性的疗方法,设计更效的药物。现在发现有些小分子可以穿越细胞作配体与变蛋白结,从而使原已失去战能力的突变蛋白逸“白质量控制”而“带伤作战”。这种小分子被称为“药物分子伴侣”,有希望成为治疗“折”的新药。 新生肽的折叠问题或蛋白质折叠问题不具有重大的科学意义,除了面提到的在学上的应用价值外,在生物工程上具有极大的应用价值。基因工程和蛋工程已经逐渐发展成为产值以数十美元计的大产业,进入21世后,还将会有大的发展。但是当前常遇到的难,是在简单的微生物细胞内引入异体DNA 后所合成的多链往往能正确折叠成为有生物活性的蛋白质而形成不溶解的包含体降解。这一“瓶颈”问题彻底决有
生肽链折
蛋白质折叠
研究蛋质折叠在理论及实践中有意义?目前有哪些关于蛋白质折叠的学说,大致
答:在现生物化学、分子生物理学领域中,蛋白折叠/解折叠问题是引起人们极大兴趣的一个研究题,它也是生命科学领
(一)意义
蛋白质叠的研究,比较狭义定义就是研究蛋白质特定三维空间结构形成的律、稳定性和与其生物活性关系;在概念上有力学的问题和动力学的问题,有蛋白质在体外折叠和细胞内叠的问题,有理论研究和实验研究的问题。里最根的学问就是多肽链的一级结构到底如何决定它的空间结构?既然前者决定后者,一级结构和空间结构之间肯定存在某种确定的关系,有人把这设想的级结构决定空结构密码叫作“第二传密码”。蛋白质折叠机制的明将揭示生命体内的第二套遗传码,这是它的理
蛋白质分在行使生物功能时,必须具特定的二维空间结构。根据Anfinsen 理论,蛋白质分的一构决定其高级结构。蛋白质折问题就是研究蛋白质天然结是如何形的,即具有一定氨基序列的多肽链如何逐步形成蛋白质所有的空间结。目前许多“构象病”或称“折病”都是由白质折叠常造成分子集至沉引,例如,老年痴呆症(蛋白质构象变化) 、囊性纤维病变(蛋白质不能折叠) 、家族性高胆固醇症(蛋白质误折) 、家族性淀粉样蛋白症(蛋白质沉淀) 等,因此深入理解蛋白折叠机制这一需求已变得更迫切。随着蛋质折叠究的深入,人们会发现更多疾病的真正因和更针对性的治疗方法,设计更有效的药物。这是蛋白质折叠研究的
同时,它还存在重要
包涵体复性——利用DNA 重组技可以将外源基因导入宿胞。但重组基因的表达产物往往形成无活性的、不溶解的包涵。折机制的阐明对包涵体
人工设计蛋白——DNA 重组和多肽合技术的展使我们能够按照自的愿设计较长的多肽链。但由于我们无法了解这一多肽将折叠为何种构象,从无法照自己意愿
寻找致机理——许多病,如阿兹海默症(Alzheimer's)、疯牛病(Mad Cow, BSE)、可传播性海绵状脑病(CJD)、肌萎性脊髓侧索硬化症(ALS)、有帕金症(Parkinson's)等正是由于一些细胞内的重要蛋白发生突变,导致蛋白质聚沉或错误折叠造成的。因此,深入了解蛋白质折叠与错折的关系对于这些疾病的病机制的阐明以及治疗方的寻找将大
揭示蛋质功能——基因组序的发展使我们得到了大量的蛋白质序列,结构息的获得对于揭示它们的生物功能是十分重要的。依靠现有手段(X-ray 晶体衍射、NMR 及电)测定白质的结构需要较长的时间,因此结构解析步伐已发现蛋白的步伐。而结构预测的方法虽然速度较快,但可靠性并不高,只有当我们对于维持蛋白质结构,驱动蛋白质折叠的理化因更为了解,这一法才可能有根的改进。另外,对蛋白相互作用、配体与蛋白质的用等结构与功能关系的研究也有赖蛋白质折叠机制
(二)学说
1.Anfinsen 的“自组装热力学假说”
由Anfinsen 等根据对RNase 复性研究的典实验提出的经典的“热力学假说”为,天然蛋白质多肽链采取的构在一定环境条件下热力学上最稳定的结果,采取天然构象的多肽链和它处的一环境条件(如溶组分、PH 、温、离子强度等)
由能最低,以处于变性状态的多肽链一定环境条件下能够自叠成天然构象。许多蛋白(特别是一些小蛋白)在体外可以可逆的行变、复性,使“热力学假
2. Ellis的“辅助性组装学说”
体内蛋白的折叠往往需要有其他助因子的参与,并伴随有ATP 的水解。因此,Ellis 1987年提出了蛋白质叠的“辅助性组装学说”。这表明白质的折叠不仅是一个热力学的过程,显然也受到力学的控。“辅助性组装学说”认,蛋白质多肽链的正确叠和组装非能自发成,在相当多的情况下是需要其他蛋白质分子的帮助,这类帮助蛋白包括分子伴侣与折叠酶。新的观点在实质上并不和Anfinsen 理论相矛,属于蛋白质折叠径或折叠的识别和装问上的认识的善,为示生理或病理条件下蛋白质的折叠理提供了新的研究思路,因而具有重要理论意义和潜在的
人们现已分离到一能在动力学上促进多肽链正确折叠的辅助因子。如分伴侣可通过与展多肽链结合而帮助多肽链进行正确非共组装;蛋白质二硫键异构酶及脯氨酸异构酶可促进具有错配二硫键的多肽链进行二硫键重排;脯氨酸顺反异构酶还可促进多肽链入正确的折途径。这些事实有力的明动力学控制在多肽正确折叠过程中所起的要作用及真
对不同的白质它们的折叠并不千篇一律,而是各有特点。从总体上讲,蛋白质折是遵循“热力学假说”的,高能态向低能态转变,但在这过程中会受到动力学上的控制。热力学控制与力学控制蛋白多肽链的折叠反应中是统一的,不同的蛋质的折过中所体出来的二者所起作用大小可能有所不同。对一些小分子单结构域的蛋白质来说,折叠过程简单,在热力学控制下较易完成;而一些结较复杂的蛋白质特是一些在折叠需要二键重排,氨酰反异构化的蛋白质在折叠过程中总体上是受热力学控制,但折叠途受动力学控制就
3. 关于分子伴侣
分子伴可以定义为“一在序列上没有相关性但有共同功能的蛋白质,它们在细胞内帮其他含多肽的结完成正确的组装,而且在组装完毕后与之分,不构这些蛋白质结构执行功能时的组份”,如休蛋白。还有一类“分子内伴侣”前肽常位于信号肽与成熟多肽之间,是成熟多肽正确折叠所必需的。折叠酶则催化蛋白质折过程共价键异构化,主有有PDI 和PPI 。有辅助折叠分子即可以是折叠又可以是分
分子伴侣在蛋白正确折叠中都起着关键性作用。首先,它可节性地阻多肽链集聚;其次, 分子伴还有折叠互补功能,即可使未形成蛋白质集聚体的蛋白质重新折叠和恢复其水溶性;此外,分子伴侣可以进折叠误的蛋白质降解。分子伴侣主要是蛋白质折叠的慢过
分子伴侣本身包括控制正确折叠所需的构象信,而只是止非天然态多肽链内的相互间的非正确相互作,或者说它们给处于折叠中间态的多肽链提供了更多的正确叠的机,因而它们能高折叠反应的
分子伴侣机体生理条件与病理条均有相关性。分子伴侣可以是一把“双刃剑”,既具有疫保用,在一定条件下还具有病作用。一方面,它可成为感染疾病中的免疫优抗原,激发宿主体内的体液免疫反和T 细介导的细胞免疫反应,如证实一些热休克蛋白在菌或寄生感中具有疫保护作用,甚至与肿瘤免疫有关。而且分子伴侣构成的“质控系统”可以防止蛋白质非活性产物干扰细胞的正常功能。但另一方面,分子伴侣也可导致疾病的发生。蛋白产物极细微的叠异,虽然对活影响不,却可以被“质控系统”滞留在内网,不能实现正常的转位、转运或分泌,导致疾病发生。还
性折叠子如Prion 朊病毒,甚可以介导正常蛋白的错误折叠,成为具有感染性的蛋白
蛋白质的折叠
中科学院生物物理研
导读
您道蛋白质折叠吗?这是一很新的。新到什么程度?您可上网到著名的不列百科全书网站检索下proteinfolding(即蛋白质折叠),还没有相应的
您道“白质折叠病”吗?疯牛病、老年性痴呆症、囊性病变、家族性胆固醇症、家族性淀粉样蛋白症、某些肿瘤、白内等等都是“折叠病”。就是相关蛋白质三维空间结构异常。这种三维间结构常是由于致病蛋白质分子通过分子间作用感染正常蛋质而造成的。请意,致病蛋白子与正常蛋白质分子的构成完全相同,只是空间
您道蛋白质折叠有多复杂吗?国“科学国人”曾经载文称,用当最快的计算机模拟计蛋白质折叠,要花一年!而当今最快的计算机已达到每秒几万亿甚至十几万亿次浮点运算的
对生命奥秘的索,将贯穿新世纪至新千
请认真阅读王志珍研究员的篇文章。不要害怕肽键、肽链、分伴侣这类专业名词,因为它们与您、您健康息息相关。读完这篇
研蛋白的折叠,是生命科学领域的前沿课题之一。白一种生物大分子,基本上是由20种氨基酸以肽键连接成肽链。肽链空间卷曲折叠成为特定的三维空间结构,括二级结构和三级结构二个主层次。的蛋白质由多条肽链组成,每条肽链称为亚基,亚基间又有特定的空间系,称为蛋质四级结构。所以蛋白质分子有非常特定的复杂的
通“蛋白质结构预测”破译“第二遗传密码”,蛋质究最后几个未揭示的奥秘之一。天津大学和中国科学院生物理所的科学家已经做出了优秀的研究成。他们预测,蛋白质的种类然成上万,但它们的折叠类型却只有有限的650种左。我国科学家在子伴侣和折方面有特色的研究成果,也已经赢得了国际同
外环境的变化可以导致蛋白质空间结构的破坏和生活的失,但却并不坏它的一级结构(氨基酸序列),这称为蛋白质变性。变性的蛋白质往往成为一条伸展肽链,在一定的条件下可以重折叠成有的空间结构并恢复原有的活性。对蛋白质变性作用认识是我国科学吴宪在三十首先提出的。 蛋白质异常的三维空间结构可以
疯病、老年性痴呆症、囊性纤维病变、家族性固醇症、家性淀粉样蛋白症、某些肿瘤、白内障等等是“折叠病”。造成疯牛病的Prion病蛋白可以感染常蛋白在蛋白质之间传染。研究蛋白质的折叠问不仅具有重大的科学意义,在医学和在生物工程领域具有极大的
1
五年代运用X射线衍射技术出了生命遗传物质脱氧核糖核酸(DNA)分子的三维空间结构,阐明了物遗传的分子基础,揭示这个最主要的生命活动的本质,从而开创在分子水上认识命现象的新学科———分子生物学。分子生出现是典生物学转变成近代生物的里程碑。 尽管自然界的生物种千万,生命现象繁杂纷飞,在分子水平研究生命,使我认识到各种生命现象基本原理却是高度一致的!从最简单的单细胞生物到最高等的人类,它们最基本最重要的组成物质都是蛋白质和。核酸是生物遗传信携带者,所有生物体世代相传,就是依靠核酸分子可以精复制的性质。蛋白质则是生命活动的主要担者。所有的命活动,呼吸、运动、消化??至感知、思维和学习,无一例外是依靠蛋白质来完成的。 蛋白质是一种生物大子,基本上是20种氨基酸以肽键连接成肽链。肽键连接成肽链称为白质的一级结构。不同蛋白质其肽的长不同,肽中不同氨基酸的组成和排列顺序也各不相同。肽链在空间卷曲折叠成为特定的三维空间结构,包括二级结构和三级结构二个主要层次。有蛋质由多条链组成,每条链称为亚基,亚基之间又有特定的空间关系,为蛋白的四级结构。所以白质分有非常特定的复杂的空间结构。每种白质分子都有自己特有氨基酸成和排列顺序,由这种基酸排列序决定它的特定的空结构,就是荣获诺贝尔的著名的Anfinsen原理。蛋白质分子只有处于它自己特定的三空间结构情况下,才能获得它特定的生物活;三维空结构稍有破坏,就很可致蛋白质生物活性的降低甚至丧
二世纪生物领域最重要的成就之一,是继DNA双螺旋结构的发现总出分子生物学的心法则,示命遗传信息传递的方向和途。近半个世纪以来对阐明中心法则有关问题有杰出贡献而获得诺贝尔奖的学者先后多34位。分生物学的中心则简单表达如下: 分子生物学的中心法中,DNA和核糖核酸(RNA)的复制、DNA转录RNA、RNA逆转录成DNA以及以信使RNA为模板翻译成多肽链的过程机制基本上已经阐明。但从多链折叠成白质的过程,即所“新生肽的折叠”问题,中法则至今留下的空
2
人对由于基因突变造成蛋白质分子中仅仅一个氨酸基变化就引起病的情况已有所了解,即所谓“分子病”,如地海镰刀状红血球贫血症就是因为血红蛋分子中第六位的谷氨酸突变了颉酸。现在则发现蛋白质分子的氨基酸序列没有改变,只是其结构或者构象有所改能引起疾病,那就是所谓“构象病”,或称“
大都知的疯牛病,它是由一种称为Prion的蛋白质的感染起,种蛋白质也可以感人而引起神经系统疾病。在正常机体中,Prion是正常活动所需要的蛋白质,而致病Prion与正常Prion的一级结构完相同,只是空结构不同。这一疾病的研究涉及到许多生物学的基本问题。一级结构完全相的蛋白为什么会有同的空间结构,这Anfinsen原理是否矛盾?显然这里有蛋白质的能量和
从认为蛋结构的变化来自于序列的变化,而序列的变化来自于基因的化,命息从核酸传递到蛋白。而致病Prion的信息已被诺贝尔奖获得者普鲁辛纳证明不是来自的变化,致病蛋白Prion导致正常蛋白Prion转为致病的折叠状态是通蛋白分子间的作而感染!种相互作用的本质和机制是什么?仅仅改变了折叠状态的分子又如何导致严的疾病?这些问题都不传统的概念给予满解释,因此在学界引起激烈的争论,有关研究的强度和竞争性也
由蛋质折叠异常而成分子聚集甚至沉淀或不能正常转运到位所引起的疾病还有老年性痴呆症、囊性纤维病变、家族性高胆醇症、族性淀粉样蛋白症、些肿瘤、白内障。由分子伴侣在蛋白折叠中至关重要的作用,分子侣本身的突变显然会引起蛋白质折叠异常而引起折叠病。随着蛋白质折叠研究的深入,人们会发现更多疾病的真正病因和更针对性的治疗方,设计更效的物。现在发现有小分子可以穿越细胞作为体与突变蛋白结合,从而使原失去作战力的突变蛋白逃逸“蛋质质量控制系”而“带伤作战”。这种小分子被称为“药物分子伴”,有希望成为治疗“折叠病”的新药。 新生肽的折叠问题或蛋质叠问题不仅具有重大的科学意义,除了上面提到的在医学上的应用价值外,在生物工程上具有极大的应用价。基因工和蛋白工已经逐渐发展成为产值以数十美元计大产业,进21世后,还将会有更大的发。是当前经常遇的困,是在简单的微生细胞内入异体DNA后所合成的多肽链往不能正确折叠成为有生物活性的蛋白质而形成不溶解的包含体或被降解。这一“瓶颈”问题彻底解决有待于
3
蛋质折叠研究,比较狭义的定义就是研究蛋白质特定三维空间结构形成规律、定和与其生物活性的关。在概念上有热力学的问题和动力学的问题;蛋白质在体外折叠和在细折叠的问题;有理论研究和实验研究的问。这里最根本的学问题就是多肽链的一结构到底如何决定的空间结构?既然前者决定后者,一级结构和空间结构之间肯定存在某种确定的关系,这是也像核酸通过“三联”决定氨基酸顺序样一套密码呢?人把这设想的一级结构决定空间结构的密码叫作“第
如说“三联密码”被破译而实际上已成为明码,那么破译“第二遗传密码”正是“蛋白质结构预测”从理论上直接地解决蛋白质的折叠题,这是蛋究后几个尚未揭示的奥秘之一。“蛋白质结构测”属于理论方面的热力学问题。就是根据测得蛋白质的一级序列预测由Anfinsen原理决定的特定的空间结构。蛋质氨基序列,特别是编码蛋质的核苷酸序列的测定在几乎已经成为常规技术,从互补DNA(cDNA)序列以根据“三密码”推定氨基酸序列,这些在上一世纪获得大突破的分子生物学技术,大大加速蛋白质一级结构的测。前蛋白质数据库中已经存有大约17万个蛋白的一级结构,但是测定了空间结构的蛋白大约只有1.2万,这中间许多是很相似的同源蛋白,真正同的蛋白只1000多个。随着人类基计划的胜利完成,解读了人类DNA全序列,蛋白质一级结构的数据增长必会出现爆炸的态势,而空间结构测定的速度远远滞后,因此二者之间还会形成更大距离,这就更
由蛋质分子结构本身极端复杂性决定了结构预测不可能一蹴而就。目前结构预测的方法大致可分两大类。一类是假设蛋白质分子然构象于热力学最稳定,能量低状态,考虑分子所有原子间的相互用以及蛋白质分子与溶剂之间的互作用,采用分子力学的能量极小化方法,计算出蛋白质子的天然空间结构。第二类方法是找出数据库中已有的蛋白质的空间结构与其一级序列之的联系总出一规律,逐级从级序列预测二级结构,再建立能的三维模型,根据总结出的间结构与其级序列之间的规律,排不合理的模型,根据能量最低原理得到修正的结构。这也就所谓“基知识的预测方法”。但是,第一类方法遇到数学上难以解决的多重极值题,而逐级预测又受到二级结构预测精度的限制。因此必须解决这些困难,或者发展新的方法,将基于知识的预测法与计算化以及统计理学结合起来,才有希望能破译“第二遗密码”。 另方面,和以往只能利用存入蛋白数库的数据进行预相比,人类DNA的全序列测定给蛋白质结构预测更自然的、信息量更得多的数据库,因此可用基于同源性的重复循环技术非常可靠地灵敏地进行结构预测。已经有人根基因组的数据用法重新估计了蛋白质折
型目大为1000种,这和早期的理论估计是一致的。显然,类基因全序列的示必然为蛋白质结构预测、蛋白质相互作用的预测以及核苷酸多态性的分子表型预测开辟前所未有广阔天地。天津大学和中国科院生物物所的科学家已经活跃在蛋白质结构预测领域,并做出了优的研成果。他预测,蛋白质的
蛋折叠第二根本的科学问题是具有完整一级结构的多肽链又是如何折叠成为它特的高级结构?这是个折叠的力学问题,长期以来,主要用体外的验方法研究,虽然已有四五十年,但至今尚未解决。我们知道,多数蛋白质在体外是不稳定的,外界变化,如度、酸度等,都以导致其空间结构破坏和物活性的丧失,但却并不坏它的一级结构,这称为蛋白质的性。对蛋白质变性作用的识是我国科学家在三十年代基于他在国内的工作首先提出来的,长期以来已经为国际上泛接受。变性的蛋白质往往成为一伸展肽链,于一级结构仍然完整,据Anfinsen原理应可以在一定的条件下重折叠成原有的空间结构并恢复原有的活性。这就是长时间来在体外研究蛋白
现道,绝多数蛋白质从一条伸展的肽链,折叠成有其特定结构的、有活性蛋白质,并不是一完成的,而要过许多折叠的中间状态。含有个亚基的蛋白质分子,亚基间的相互作用使之组装成复杂蛋白分子。研究人员用实验方法,别近年来发的快速测定方法追踪蛋白质重折叠的全程,尽可能捕捉折叠过中的每一个中间状态。不同阶段折叠速度不同,有的比,比较容易发捕捉;但有的非常快,必须要有特殊的设备配合各种测试技术去进研究。最近有人尝试大幅度降低度使折叠速减慢而得以追踪。,人们要定量地描述整个叠动态过程,拍出一部白质折叠的电影来,但这必然要经过一个长时间的艰苦的工作。 4
尽年来体外白质折叠研究为揭示蛋白质折叠的本质提供了大量信息,但细胞内白质的生物合成,个广义的白折叠问题,是一个比试管内蛋白折叠复杂得多的多的过程。蛋白质的多肽链都是在细胞内的一种由多种蛋白质和核糖核酸所组成的核糖体复合物上,以信使糖核酸为模板,从氨基末开始,按照三联密码,个氨基酸一个氨基酸加上去而合成来的。现在比较一致的看认为,这种新合来的多肽链(称为新生肽)在合成过程中长度不断增加,并在延伸的时也在进行着折叠,而不是在合成成离核糖后再自发折叠成为蛋。所以上面介绍的在体外变性展肽链的重折叠研究白质折叠的模型看来并不是研究细胞内蛋白质折叠的理想模型。由于每个
时带多个核糖体,而每核糖体上的多肽链的延伸程度是不同的,所以糖体上的多肽链合成同步化是目前研新生肽折叠的关键问题,但一直没有
我实验暂时绕过这个障碍,制备一系列从氨基末端开始具不同的肽段,比较研究们的构象作为模型,对新生肽在合成延伸的同时也在进行着叠的观点已经提供了大量信息。从核糖体上合出来的肽链还需经与翻译同时进的和翻译成后的化学加工,如形成二硫键,完成糖基化作用、羟基化作、磷酸作用等化修饰。化学修饰往肽链的折叠密切相关,没有化学修饰的肽链往往不能完
新肽还被运送到细胞的特定部,“各就各位”才能发挥它特定的生物功能:例如进入细胞核中的核蛋白与DNA组成染色体;进入线粒的蛋白参与能量代谢;组成膜的蛋白以及分到细胞外蛋白必须入内质网,先进行加工继续转运等等。这些都一个越结构,甚至是多次越膜过程。有完整空间结构的蛋白分子不能越的,因此在转运过程中折叠过多的分子必须解开折叠后才越膜。此外,多亚基白必须进行组装。有些蛋白质,如一些酶和激素,以前体形式合成后还要经过水解除去某一段序列后才能成熟为有活性的。所有这些都含在新成熟为功能蛋白的全程中,每一步都涉及新生肽链的构象变化、折叠和调整。 在试管中做蛋白质折叠实验条件往往人为化或不得不简化,与新生肽在细胞折叠的条件有质的或的差别。所有的细胞中都存在着大量的蛋白质、核酸、多糖等种生物大分子,们大约占用细胞容积的20-30%,总浓度高达每升80-200克,因此任何一种大分子都于一充满其他分子的“拥挤”环境中,使得任何一个大分子的实际可及空间大大减少,这种情况对所有大分子之间的反应在热力学动力学上都有很大的影响。近,有人呼吁,在体外研究蛋白折叠必须考虑模拟细胞内的“拥挤”环境。我验室在方面的研究已经得国际同行注意。此外,某一种蛋白在某时刻细内的局部浓度可以非常,这样高度的蛋白质在试管中必然生聚集而不能完成折叠。所以,体外进实验,为了提高白折叠效率,并且有利于进行分析,实验所用的蛋白浓度总是很低的;温度常在37摄氏度以下,有时低到10摄氏度以,以减缓反速度。溶液成分也尽量,便于分析。 5分子伴侣蛋白和折
最15年,由于发现一些蛋白质的折叠必须在另一些蛋白质存在才能完成的现象,对蛋白折叠的概念产生了革命性的全新认识,“自发折叠”的经典概生了转变和更新,这是蛋白质折叠研究中的大事。在认为新生肽在细胞的折叠和成熟在数情况下不能自发完成的,而是需要别的蛋白质帮助的。这个新概念并不与Anfinsen原理相,而是在动力学点上完善了Anfinsen学说。一个在热力学上可以成立
力的能障等问在实际上未必以完成,
目已识到的在细胞内帮新生肽链折叠的蛋白有二类:一类称为分子伴侣蛋白,另一是催化与折叠直接关的化学反应的酶,又称折叠酶。子伴侣然是一种具有新功能的蛋,近年来已经鉴越越新的分子伴侣蛋白或知蛋白的新的分子伴侣活性。它们精细三维结构、结构与功能的关系、它们帮助生物大分子折叠机制都在活跃的研究之中;特别是有些蛋白的分子伴侣活性和在同一分子上的其他生物活性之间关系以及在命中的协作和调控引起人们的兴趣。现在发现,不蛋白质的折叠需要分子伴侣的帮,DNA分和RNA分子的折叠也往往要分子伴侣的帮,因为功能DNA和RNA分子,特别是它们蛋白质形成复合物都要有一定的构象。DNA和RNA分本身具有较大的刚性,不容易叠在折叠过程中容易发生折叠错误,因此需要DNA分子伴侣或RNA分子伴侣帮助它们折叠而形成特定的构象。另一方面,不蛋白质,现在发现有些核酸和磷脂也能发挥分子伴侣的用;更有的是最近发现糖体有分子伴侣活性。有些生物分在成熟过程中需要系列分子伴侣在不同的阶给予帮才能完成最终的折叠。另外,有一些小分物质对某些蛋白质在体外的折叠有帮助作用,被称之为“化学分子伴侣”。我国科学家在分子伴和折叠方面的有特色的研已经赢得了国际同行的注
新肽细胞中折叠和成的过程由于转录或翻译出现了错误,或受到各种环境刺激而损,并非能百分之百完成。为提高蛋白质生物合成的效,处理不能继续正确折叠的或错折叠的“次品”“品”,防止这些“垃圾”的积而危害正常生命活动,生命的化使细胞获得了一种“蛋白质质量控制系统”。这种系统是由子伴侣和靠消耗三磷酸腺苷的能量而发挥作用的特定的蛋白水解酶组成。分子伴侣帮助新生肽正折叠;而特的蛋水解酶把不能继续确折叠的或错误折叠的“垃圾”水解成小分子。这里的科学问题是量控制系统底如何进行质量控制?有借用了医院里诊病人应该送到哪种病房作什么治疗的机制(triage)描述细胞的蛋白质质量控制体系的作用机理。键是如何“诊断”和区分什样的生肽“病人”可以送到分子伴侣“病房”进行治疗和拯救,而什么样的新生肽“病人”已“无可救药”,只能送给蛋白水酶处理。胞内新生“病人”的命运主要是由分子伴侣处“病人”的能力和速度动力学来决定的。尚未治好的新生“人”可能获得再治的机,但也可能不幸送到白水解那里去了;还有一种可能性就是形成聚,聚集体是抗水解的。如果形成有规则的聚集体,即所谓“淀粉样纤维”。一些神经系统退化疾,如老性痴呆症、帕金森、亨廷顿氏病就是由此造成
在21世纪,人类在解决了新肽折叠的问题,解决了基因调控问题后,应该就可说全面地最终地阐了分子生物学的中心法,那时人类对自身的认识又将有一个新的
作者简介
王珍,女,中国科学院生物物理研究所生物大分国点实验室研究、博士生导师。曾在胰岛素和胰岛素受体的结构功能关系以及胰岛素作用机制方面做研究。近年来主要从事蛋白折叠、叠酶和分子伴侣方面的研究。曾获国家自然科学二奖,中国科学自然科学一奖、二等奖等,获“国家有突出贡献的中青年专